Библиотека
Теология
Конфессии
Иностранные языки
Другие проекты
|
Ваш комментарий о книге
Лоренц К. Оборотная сторона зеркала
Глава 2 ВОЗНИКНОВЕНИЕ НОВЫХ СИСТЕМНЫХ СВОЙСТВ
1. НЕДОСТАТОЧНОСТЬ СЛОВАРЯ
Когда мы пытаемся изобразить великий процесс становления жизни в согласии с его природой, мы неизменно сталкиваемся с тем препятствием, что словарный запас культурных языков сложился в то время, когда единственным известным видом развития был онтогенез, т. е.
индивидуальное развитие живого существа. В самом деле, такие слова, как Entwicklung'*, Development2*, Evolution3* и т. п., все означают в этимологическом смысле, что развивается нечто, уже бывшее прежде в неразвитом или свернутом состоянии, подобно цветку внутри почки или цыпленку внутри яйца. Указанные выражения удовлетворительно описывают такие онтогенетические процессы. Но они, к несчастью, полностью отказываются служить, когда мы пытаемся правильно изобразить сущность творческого процесса, состоящего именно в том, что все время возникает нечто совершенно новое, чего прежде попросту не было. Даже прекрасное немецкое слово Schopfung (творение)4* этимологически означает, что нечто уже существующее черпается из некоторого также существующего резервуара. Некоторые философы-эволюционисты, осознав недостаточность всех этих слов, ухватились за еще худшее слово "эмергенция"*, вызывающее по логике языка представление о чем-то заранее сформировавшемся и внезапно появившемся, подобно киту, вынырнувшему для вдоха на поверхность моря, которое при буквально поверхностном рассмотрении казалось пустым.
2. ФУЛЬГУРАЦИЯ
Философы-теисты и мистики средневековья ввели для акта сотворения нового выражение "Fulguratio", что означает вспышку молнии. Несомненно, они хотели выразить этим непосредственное воздействие свыше, исходящее от Бога. По этимологической случайности — если не вследствие более глубоких неожиданных связей — этот термин гораздо лучше приведенных выше выражений описывает процесс вступления-в-существование чего-то прежде не бывшего. Для естествоиспытателя Зевсов перун — такая же электрическая искра, как и всякая другая, и если мы замечаем искру, проскочившую в неожиданном месте системы, то первое, что нам приходит на ум, это короткое замыкание — вновь возникшая связь.
Когда, например, совместно включаются две независимые системы, как это изображено на приведенной рядом простой электрической модели, заимствованной из книги Бернгарда Гассенштейна, то при этом внезапно возникают совершенно новые системные свойства, ранее не существовавшие даже в зачаточном виде. Это и есть глубокое содержание мистически звучащего, но вполне справедливого принципа гештальтпсихологии: "Целое больше своих частей".
Особый случай возникновения новых системных свойств (ряд примеров которого нам еще встретится в дальнейшем) состоит в следующем. В последовательности подсистем, соединенных друг с другом в линей-
1 *Развитие (нем.). Подобно русскому слову и приводимым дальше словам, означает развертывание чего-то свернутого (от wickeln — мотать, навивать, обертывать). — Примеч. пер.
2Развитие (англ.). — Примеч. пер.
3 *Развитие, эволюция (от лат. evolutio — развертывание (свитка)). — Примеч. пер.
4*Schopfung от schopfen — черпать; Schopfung означает сотворение мира и все мироздание, но Schopfwerk — водокачка. — Примеч. пер.
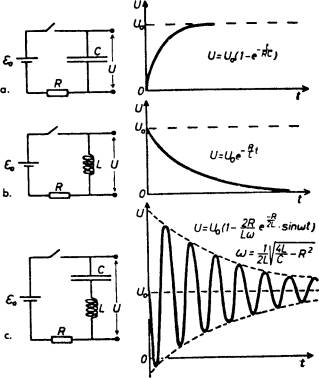
Рис. 1. Три электрические цепи, в том числе колебательный контур (с), иллюстрирующие понятие "системное свойство". Полюсы батареи с электродвижущей силой E и напряжением на зажимах U ц соединены проводником. Омическое сопротивление цепи в целом обозначается Я. В случае (а) в цепь включен конденсатор емкости С, в случае (b) — катушка индуктивности (-, в случае (с) — и конденсатор, и катушка. Напряжение U может быть измерено на двух зажимах. Графики справа показывают изменение напряжения после включения тока в нулевой момент. В случае (а) конденсатор постепенно заряжается через сопротивление до тех пор, пока он не достигает напряжения U В случае (b) сила тока — вначале сдерживаемая самоиндукцией — возрастает до тех пор, пока не достигнет заданной законом Ома величины; при этом напряжение U теоретически обращается в нуль, поскольку общее сопротивление цепи заключено в Я. В случае (с) возникают затухающие колебания. Наглядно очевидно, что поведение системы (с) не может быть получено простым сложением процессов (а) и (b), хотя можно представить себе, что (с) получается соединением (а) и (b).
Схема действует, например, при следующих значениях величин: С = = 0,7 · Ю-9 F: L = 2 · 103 Ну; R = 103 ?; ? » 1,2 •??-6.
Последнее значение определяет также общую для всех трех кривых временную ось. (Расчет выполнил Э. У. фон Вейцзеккер.)
ную цепь, где каждая подсистема причинно воздействует на следующую, так что первая может быть только причиной действия, а последняя функционирует только как действие, возникает новая причинная связь, с помощью которой· именно эта последняя система приобретает влияние на первую; вследствие этого причинная цепь замыкается, превращаясь в круг. С примерами таких замкнутых цепей, а именно цепей с положительной обратной связью, мы уже познакомились при рассмотрении способов получения энергии и информации. Не менее важное значение имеют круговые процессы с отрицательной обратной связью; но, поскольку они относятся к механизмам получения информации, я займусь ими подробнее в соответствующем разделе. Здесь же достаточно указать следующее. Когда в круг причинных связей встроен в некотором месте "знак минус" и тем самым действие процесса уменьшается в этом месте цепи тем больше, чем сильнее действие в предыдущем звене, то отсюда возникает эффект регулирования. Например, чем выше уровень жидкости в резервуаре карбюратора или в туалетном бачке, тем выше поджимается поплавок, запирая тем самым дальнейший приток жидкости. Следствием этого процесса является постоянство уровня жидкости.
Кибернетика и теория систем, объяснив внезапное возникновение новых системных свойств и новых функций, устранили неприятную видимость, будто в таких случаях происходят чудеса. Нет совершенно ничего сверхъестественного в том, что линейная причинная цепь замыкается в круг и тем самым возникает система, отличающаяся от всех предыдущих вовсе не степенью ее свойств, а принципиально. "Fulguratio" этого рода может быть единственным по значению, в самом подлинном смысле слова эпохальным событием в эволюции.
3. ВОЗНИКНОВЕНИЕ ЕДИНСТВА ИЗ МНОГООБРАЗИЯ
Как поняли многие мыслители, и философы, и естествоиспытатели, прогресс в развитии жизни почти всегда достигается таким образом, что некоторое число различных и ранее функционировавших независимо друг от друга систем интегрируется в некоторое целое высшего порядка и что в процессе такой интеграции в них происходят изменения, делающие их более подходящими для сотрудничества со вновь возникающей целой системой. Как известно, Гёте определил развитие как дифференциацию и подчинение частей. Людвиг фон Берталанфи с большой точностью описал этот процесс в своей теоретической биологии и привел ряд примеров. У. Г. Торп весьма убедительно показал в своей книге "Наука, человек и нравственность", что возникновение некоторого целого из многообразия различных частей, которые становятся при этом еще более непохожими друг на друга, есть важнейший творческий принцип эволюции: "Unity out of diversity"'*. Наконец, Тейяр де Шарден выразил то же в самой краткой и поэтически самой прекрасной форме: "Creer, c'est unir"2*. Этот принцип, по-видимому, действовал уже при самом возникновении жизни.
1 *"Единство из многообразия" (англ.). — Примеч. пер.
2 ""'Творить — значит соединять" (фр.). — Примеч. пер.
Творческое соединение многообразного в единое функциональное целое само по себе означает усложнение живой системы. Но в ходе дальнейшей эволюции новая система часто упрощается посредством "специализации" соединившихся в нее подсистем; именно, каждая из них ограничивается той функцией, которая приходится на ее долю при новом разделении труда, тогда как другие функции, которые ей также приходилось выполнять во время ее независимости, переходят к другим членам целой системы. Даже клетки ганглий* нашего мозга, осуществляющие в совокупности высшие духовные функции, каждая в отдельности далеко уступают амебе или туфельке, причем не только в отношении отдельной функции клетки, но также и в отношении существенной информации, лежащей в основе этой функции. Амеба или парамеция располагает целым рядом осмысленных ответов на внешние стимулы и "знает" много важных вещей об окружающем мире. Но клетка ганглии "знает" лишь, когда она должна выстрелить, и даже этот выстрел она не может сделать ни сильнее, ни слабее: он должен быть лишь произведен или нет по принципу "все или ничего". Это "поглупение" члена, встроенного в высшее целое, имеет, естественно, свой положительный смысл: оно необходимо для функции целого, поскольку обеспечивает однозначность передачи сообщений. "Депеша", переданная клеткой, не должна быть слабее или сильнее в зависимости от случайного, сиюминутного состояния клетки, точно так же как дисциплинированный солдат не должен выполнять приказ с большей или меньшей энергией по его собственному усмотрению.
Такое упрощение первоначально независимых подсистем в ходе их интеграции в высшее целое представляет собой явление, наблюдаемое на всех ступенях эволюции. На уровне психосоциального развития человека и его культуры оно ставит перед нами трудные проблемы. Неизбежное развитие обусловленного культурой разделения труда неудержимо ведет к возрастающей специализации во всех человеческих занятиях и хуже всего в науке. Процесс этот приводит к тому, что, как говорит старая острота, специалист знает все больше и больше о все меньшем и меньшем и в конечном счете знает все ни о чем. Существует серьезная опасность, что специалист, которого конкуренция коллег вынуждает ко все более детальному и более специальному знанию, будет все меньше и меньше ориентироваться в других отраслях знания, пока наконец не утратит всякое суждение о том, каково значение и роль его собственной области в рамках большей системы отсчета, сверхличного общего знания человечества, составляющего достояние его культуры. В следующем томе мне придется вернуться к проблеме специалиста*.
Другой вид упрощения более высокоорганизованной системы составляет то, что в человеческой общественной жизни называется "улучшением организации". Так же как любая сконструированная человеком машина в своих первых, опытных образцах устроена сложнее, чем в окончательном варианте, часто упрощаются и живые системы. Взаимодействия, в частности обмен информацией между подсистемами, упрощаются или направляются на более прямые пути, ненужные исторические пережитки устраняются или, как это называется на биологи-
ческом языке, "рудиментируются". Особенно типично упрощение посредством "лучшей организации", происходящее в сверхличных, объединенных культурой сообществах людей.
4. ОДНОСТОРОННЕЕ ОТНОШЕНИЕ МЕЖДУ УРОВНЯМИ ИНТЕГРАЦИИ
Описанный выше способ интегрирования уже существующих подсистем в некоторое функциональное целое имеет своим следствием весьма своеобразное, в известном смысле одностороннее отношение, равным образом существующее внутри организма, между его целой системой и подсистемами, а также между высшими организмами и их уже вымершими примитивными предками. То же отношение существует в принципе между всем живым и неорганической материей, из которой оно состоит. Это отношение можно выразить онтологически* словами: целое есть все его части и продолжает этим оставаться, даже если в процессе эволюции оно обогащается вдобавок рядом новых системных свойств, создаваемых последовательными "фульгурациями". Сами подсистемы не приобретают при этом никаких новых и высших системных свойств, более того, в процессе уже описанного упрощения они могут даже потерять некоторые из них. Но ни одна закономерность, присущая подсистемам, не нарушается их вхождением в целое, и меньше всего — закономерности, управляющие неорганической материей, из которой построено все живое.
Таким образом — ив этом состоит рассматриваемый здесь односторонний характер отношения, — целая система обладает всеми свойствами своих членов, и прежде всего разделяет все их слабости, поскольку никакая цепь, разумеется, не может быть прочнее ее слабейшего звена. Но ни одна из множества подсистем не обладает свойствами целого. Весьма сходным образом каждый высший организм обладает большею частью свойств своих предков, но в то же время никакое сколь угодно точное знание свойств некоторого живого существа не позволяет нам предсказать свойства его выше развившихся потомков. Это вовсе не означает, что высшие системы не поддаются анализу и естественному объяснению. Но при этом исследователь в своих аналитических устремлениях никогда не должен забывать, что свойства и закономерности всей системы в целом, а также любой из ее подсистем всегда должны объясняться, исходя из свойств и закономерностей тех подсистем, которые находятся на ближайшем, низшем уровне интеграции. А это возможно лишь в том случае, если известна структура, соединяющая подсистемы этого уровня в высшее единство. Предполагая полное знание такой структуры, можно в принципе объяснить естественным способом, т. е. без привлечения сверхъестественных факторов, любую, даже самую высокоорганизованную живую систему со всеми ее функциями.
5. НЕ ПОДДАЮЩИЙСЯ РАЦИОНАЛИЗАЦИИ ОСТАТОК
Впрочем, это утверждение о принципиальной объяснимости живого существа справедливо лишь в той мере, в какой мы считаем данными нынешние структуры его тела; иначе говоря, мы поступаем при этом
таким образом, как будто его историческое возникновение нас не интересует. Но как только мы задаем себе вопрос, почему определенный организм имеет данную, а не иную структуру, мы вынуждены искать важнейшие ответы в предыстории соответствующего вида. Если мы спрашиваем, почему наши уши находятся как раз на этом месте — по обе стороны головы, то этот вопрос допускает законный каузальный* ответ: потому что мы происходим от предков, дышавших в воде и имевших в этом месте жаберную щель, так называемое брызгальце, сохранившееся при переходе к сухопутному образу жизни в качестве проводящего воздух канала и после изменения функции используемое для слуха.
Число чисто исторических причин, которые надо было бы знать, чтобы объяснить до конца, почему организм устроен "так, а не иначе", если не бесконечно, то, во всяком случае, настолько велико, что для человека в принципе невозможно проследить все такие цепи причинных связей, даже если бы они имели конец. Таким образом, всегда остается, как говорит Макс Гартман, некоторый иррациональный, или не поддающийся рационализации, остаток. То обстоятельство, что эволюция произвела в Старом Свете дубы и человека, а в Австралии — эвкалипты и кенгуру, обусловлено именно этими уже не поддающимися исследованию причинами, которые мы обозначаем обычно пессимистическим термином "случай".
Хотя, как надо снова и снова подчеркивать, в качестве естествоиспытателей мы не верим в чудеса, т. е. в нарушения всеобщих законов природы, мы вполне отдаем себе отчет в том, что нам никогда не удастся до конца объяснить возникновение высших живых существ из их более низкоорганизованных предков. Как подчеркнул в особенности Майкл Поланьи, высшее живое существо не "сводимо" к своим более простым предкам, и тем менее живая система может быть "сведена" к неорганической материи и происходящим в ней процессам. Впрочем, в точности то же относится к сделанным человеком машинам, которые представляют поэтому хорошую иллюстрацию того, в каком смысле здесь понимается несводимость. Если имеется в виду их современное, физическое устройство, то они до конца поддаются анализу, вплоть до идеального доказательства правильности анализа — полной осуществимости синтеза, т. е. практического изготовления. Но если имеется в виду их историческое, телеономное* становление как органов Homo sapiens, то при попытке объяснить, почему эти машины устроены "так, а не иначе", мы сталкиваемся с таким же не поддающимся рационализации остатком, как и в случае живых систем.
Как можно предположить, Поланьи далек от того, чтобы постулировать виталистические факторы; но, чтобы полностью исключить такое недоразумение, я предпочитаю говорить, что система, принадлежащая более высокому уровню интеграции, не выводима из более низкой, как бы точно мы ее ни знали. Мы знаем с полной уверенностью, что высшие системы возникли из низших, что они построены из них и до сих пор содержат их в качестве составных частей. Мы знаем также с полной уверенностью, каковы были предыдущие стадии, из которых возникли
высшие организмы. Но каждый акт построения представлял собой "Fulguratio", случившуюся в эволюции как единственное, своеобразное событие, и это событие в каждом случае носило характер случайности или, если угодно, изобретения.
Глава 3 СЛОИ РЕАЛЬНОГО БЫТИЯ
1. КАТЕГОРИИ БЫТИЯ НИКОЛАЯ ГАРТМАНА*
"Справедлив ли известный упрек в адрес философии Канта, что из нее вовсе исключен вопрос об основе бытия? Не следует ли, напротив, полагать, что проблема проведения границ и вообще проблема объективной истинности как раз и ставит вопрос об основах бытия? В самом деле, понятие лишь в том случае может подходить к некоторому предмету, если оно приписывает ему качества, действительно ему присущие. Таким образом, "объективная истинность", во всем ее объеме, предполагает, что категория разума есть в то же время предметная категория".
Как видно из этих высказываний Николая Гартмана, то, что побуждает его отождествить категории человеческого мышления с категориями внесубъективной реальности, — это его глубокое, коренное убеждение в существовании подлинного внесубъективного мира. Категория означает для него высказывание, предикат. Он говорит: "Категории суть основные предикаты сущего, предшествующие всем специальным соотношениям (Predikationen) и в то же время образующие их рамки". И в другом месте: "... сами они суть наиболее общие формы высказываний — а также колеи для возможных более специальных высказываний, — и тем не менее они выражают основное строение предметов, к которым они относятся. Смысл этого состоит в том, что выраженное таким образом основное строение предметов как раз присуще им в их бытии, и притом независимо от того, высказаны эти суждения о предметах или нет (курсив мой. — К. Л.}. Все сущее, когда оно высказано, является в виде предикатов. Но предикаты не тождественны с ним. Понятия и суждения создаются не ради них самих, а ради сущего.
Внутренний, онтологический смысл суждения в том, что его логически имманентная форма трансцендирует*. Именно это придало понятию "категория" его онтологическую устойчивость, вопреки всем недоразумениям".
Если Гартман, как это достаточно ясно вытекает из цитированных утверждений, уверенно предполагает, что категории разума суть в то же время категории предметного мира, и если он основывает на этом, как он в действительности делает, свое убеждение в существовании и в относительной познаваемости внешнего мира, то он тем самым ближе всего подходит в своих основных гносеологических позициях к точке зрения гипотетического реализма, для которого категории и формы созерцания человеческого познавательного аппарата представляются.
чем-то очевидным образом возникшим в ходе эволюции и "подходящим" к условиям внесубъективного мира — аналогичным образом и по аналогичным причинам, как лошадиное копыто "подходит" к поверхности степи или плавник рыбы к воде. Конечно, Гартман был весьма далек от того, чтобы искать генетически-историческое объяснение утверждаемого им соответствия между категориями разума и категориями предметного мира. Однако его взгляды на строение реального мира, в особенности мира живых организмов, развитые им на основе его учения о категориях, столь полно совпадают со взглядами филогенетика*, что я всегда затрудняюсь пересказать ход мыслей Гартмана, не протаскивая в его учение о слоях эволюционные интерпретации. Однажды я спросил моего друга Вальтера Роберта Корти, близко знавшего Гартмана, что, по его мнению, сказал бы великий философ об эволюционном истолковании его учения. Корти ответил, что Гартман отверг бы такое истолкование, но прибавил утешительно: "Впрочем, именно это делает его теорию съедобной"*. В следующем разделе я опираюсь на это изречение подлинного философа.
2. УЧЕНИЕ НИКОЛАЯ ГАРТМАНА О СЛОЯХ РЕАЛЬНОГО БЫТИЯ
В реальном мире, где мы живем, говорит Николай Гартман, мы находим слои, каждому из которых присущи особые категории бытия или группы категорий бытия, присутствие или отсутствие которых отделяет этот слой от других. "В ступенчатом устройстве реальности имеются определенные основные феномены, отделенные непреодолимыми различиями", так что "согласованная с феноменами теория категорий должна принимать во внимание эти подразделения, так же как нарушающие их связи между явлениями бытия..." Но эти связи, проникающие через границы, которыми отделены друг от друга четыре великих слоя реального бытия — неорганического, органического, психического и духовного — действуют всегда односторонне. Принципы бытия и законы природы, справедливые в неорганическом мире, сохраняют свою безусловную силу и в высших слоях. Гартман пишет: "Таким образом, органическая природа возвышается над неорганической. Она не может, однако, быть независимой, а предполагает отношения и закономерности материального мира; она основывается на них, хотя они никоим образом не достаточны, чтобы составить живое. Точно так же психическое бытие и сознание обусловлено несущим их организмом, в котором и вместе с которым они только и могут явиться на свет. Подобным же образом великие исторические явления духовной жизни связаны с психической жизнью индивидов, их носителей в тех или иных условиях. Перемещаясь от слоя к слою через разделяющие их границы, мы каждый раз обнаруживаем одно и то же отношение основанности на "низшем", обусловленности этим "низшим" и в то же время самостоятельности высшего в его своеобразии и собственной закономерности.
Это отношение и есть, по существу, единство реального мира. При всем разнообразии и всей неоднородности мира он вовсе не лишен единства. Он обладает единством некоторой системы, но это система, состоящая из слоев. Реальный мир имеет слоистое строение. Дело здесь
не в том, что разделительные границы непреодолимы — потому что они, может быть, непреодолимы только "для нас", — а в появлении новых закономерностей, новых способов образования категорий, хотя и зависящих от низшего слоя, но своеобразных и самостоятельных по отношению к нему".
Это прекрасное место из работы Гартмана свидетельствует, таким образом, о фундаментальном совпадении его чисто онтологически обоснованных взглядов со взглядами филогенетика, черпающего свое знание из сравнительного и аналитического изучения живых организмов. Учение Гартмана о слоях осуждалось как "псевдометафизическая конструкция", и совершенно несправедливо, потому что оно вовсе не является таковой. Оно построено не на дедуктивной спекуляции, а на эмпирическом материале и согласуется с явлениями и многообразием нашего мира, не разрывая его на разнородные составные части.
С моей точки зрения, самое убедительное доказательство онтологической правильности этого учения состоит в том, что оно, вовсе не принимая во внимание факты эволюции, тем не менее в точности с ними согласуется, подобно тому как согласовалась с ними любая хорошая работа по сравнительной анатомии, даже выполненная до Дарвиновых открытий. Построенная Гартманом последовательность великих категорий бытия просто-напросто совпадает с порядком их возникновения в истории Земли. Неорганическое существовало на Земле очень долго до появления органического; гораздо позже в ходе эволюции возникали центральные нервные системы, которым можно приписать субъективное переживание, т. е. "психику"*. И наконец, лишь в новейшей стадии творения выступила на сцену духовная жизнь.
Гартман отчетливо говорит, что категориальные различия между низшими и высшими слоями никоим образом не сводятся к великим границам, разделяющим неорганическое и органическое, органическое и психическое, наконец, психическое и духовное. Он говорит: "Высшие формации, из которых состоит мир, расслоены подобно тому, как расслоен сам мир". Для нас это означает, что каждый шаг эволюции, ведущий от существа низшей ступени организации к существу высшей ступени, в принципе имеет тот же характер, что и само возникновение жизни.
3. НАРУШЕНИЯ ПРАВИЛ КАТЕГОРИАЛЬНОГО АНАЛИЗА ФЕНОМЕНОВ И КАУЗАЛЬНОГО АНАЛИЗА СИСТЕМ*
Согласованность онтологии Гартмана с эволюционным изучением систем становится особенно очевидной, если рассмотреть, наряду с их закономерными методами, также их, к сожалению, столь частые нарушения. Онтолог озабочен тем, чтобы его описание внешней действительности согласовалось с феноменами, т. е. чтобы никакому явлению действительности не приписывались не подходящие к нему категории бытия, и при этом не были бы пропущены никакие категории, которые для этого явления характерны. Гартман говорил: "Легко усмотреть, что в устройстве реального мира существует некоторое расслоение; непредубежденный взгляд просто неизбежно обнаруживает его. И оно в самом.
деле было замечено уже давно. Но представление о расслоенности не могло быть беспрепятственно принято по той причине, что ему всюду противостоял постулат единства мира. выдвинутый спекулятивным мышлением ".
Метафизическая спекуляция проявляется, например, в том, каким образом радикальный механицизм пытается объяснить все мироздание категориями и закономерностями классической механики, попросту недостаточными для этой цели. Когда механицист пренебрегает при этом закономерностями высшего порядка, отделяющими более высокие слои от низших и поднимающими их над низшими, или вовсе их отрицает, то возникает очевидная, но прямо-таки неискоренимая ошибка перехода границы "снизу вверх". Все так называемые "измы", такие, как механицизм, биологизм, психологизм и т. п., стремятся охватить процессы и закономерности, характерные и единственно свойственные высшим слоям, категориями явлений, относящимися к низшим, что попросту невозможно.
Такое же насилие над наблюдаемыми явлениями происходит, когда разделительная граница нарушается в обратном направлении. Гартман говорит об этом заблуждении, в некотором смысле противоположном предыдущему: "Исходный пункт всей картины мира выбирается в этом случае на высоте психического бытия — где человек переживает ее в своем самоощущении, — и оттуда принципы переносятся "сверху вниз", на низшие слои реальности". Все панпсихические картины мира, как, например, учение Лейбница о монадах, учение Якоба фон Юкскюля об окружающем мире и даже остроумная попытка Вейделя разрешить психофизическую проблему, впадают в одну и ту же ошибку, пытаясь объяснить все многообразие мира на основе единственного рода принципов бытия и становления.
Потребность в таком образе действий с целью прийти, насколько возможно, к единой картине мира у многих мыслителей кажется прямо непреодолимой. Иначе невозможно объяснить, как может человек со здравым рассудком отказывать собаке или шимпанзе в субъективном переживании, как это делал Декарт, или приписывать такие свойства атому железа, чему показал пример Вейдель.
Все открытия современной системной теории эволюции, относящиеся к возникновению новых системных свойств и к одностороннему отношению между разными уровнями интеграции, отчетливо свидетельствуют, что каузальный анализ, согласный с системными свойствами живой системы, приводит к результатам и связан с методами, которые весьма близки к результатам и методам гартмановского согласованного с феноменами категориального анализа. Более того, можно даже утверждать, что лишь согласованный с системами каузальный анализ позволяет показать, почему порицаемые Гартманом нарушения границ приводят к столь тяжким заблуждениям. Мы в точности понимаем, почему невозможно вывести свойства более высокоинтегрированной системы из свойств низшей системы (см. с. 275), а также почему столь бессмысленно пытаться проследить — и тем более постулировать — в отдельных подсистемах целой системы или у более простых предков высшего организма те свойства и функции, которые начинаются лишь с творческого акта высшей интеграции.
В этой книге мне придется особенно много заниматься одним типичным заблуждением этого рода — упорным стремлением многих психологов и исследователей поведения обнаружить адаптивное обучение не только у -низших организмов, которым его попросту "еще" недостает, но, что хуже, в таких подсистемах высших организмов, которые не только не поддаются модификации посредством обучения, но, более того, по уже указанным в главе I причинам (с. 267) предохраняются от любой модификации их филогенетическим программированием. Психологу без биологической подготовки, обязанному большей частью своего практического знания о живых существах обращению с людьми и высшими млекопитающими и, сверх того, воспитанному в доктрине, рассматривающей рефлекс и условный рефлекс как простейшие и примитивнейшие элементы любого поведения животных и человека, может показаться едва ли не само собою разумеющимся приписать даже простейшим и низшим беспозвоночным хотя бы простые задатки или "орименты"* условных реакций, и он будет держаться этого заблуждения со всей страстностью, происходящей от потребности в единой картине мира. Силой этой мотивации объясняется также целый ряд самообманов — иногда поистине трагических — при попытках обнаружить "обучение" также у низших организмов.
4. ЗАБЛУЖДЕНИЕ, СОСТОЯЩЕЕ В ПОСТРОЕНИИ АНТАГОНИСТИЧЕСКИХ ПОНЯТИЙ
Как было показано в предыдущем разделе, разным слоям реального бытия соответствуют весьма различные категориальные свойства, зависящие от различных уровней интеграции системы. Были рассмотрены ошибки, происходящие от стремления понять мир из единого объяснительного принципа, объяснить более простые системы на основе слишком высоких, и обратно — высокоинтегрированные системы на основе слишком элементарных принципов.
Теперь мы займемся противоположным заблуждением человеческой познавательной деятельности, состоящим в забвении того общего, что в равной мере присуще всем слоям реального мира. Образование антагонистических понятий, противопоставление альфы и не-альфы, есть форма мышления, которая, как и стремление к объяснению из единого принципа, несомненно является врожденным свойством человека, в некотором смысле уравновешивающим это стремление.
Уже описанное в разделе 4 главы 2 (см. с. 274) одностороннее проникновение слоев снизу вверх допускает два вида высказываний об общем и о различном. Можно, например, говорить, что все жизненные процессы суть химические и физические явления, что все субъективные процессы нашего переживания суть органические, физиологические и тем самым также химико-физические процессы и, наконец, что вся духовная жизнь человека в том же смысле происходит во всех этих лежащих в основе слоях. Столь же правильно и законно говорить: жизненные процессы "по существу", т. е. в смысле принципов бытия и становления, присущих только им одним, составляющих только их существенную
особенность и прежде всего возвышающих процессы жизни над другими химико-физическими явлениями, представляют собой нечто совсем иное, чем эти явления. Сопровождаемые переживаниями нервные процессы суть нечто совершенно иное, чем неодушевленные нервно-физиологические процессы; и человек, обладающий в качестве наделенного духовной жизнью существа культурно обусловленным сверхличным знанием, умением и волей, тем самым отличается по своей сущности от своих ближайших зоологических родичей.
Между этими двумя рядами высказываний есть лишь кажущееся противоречие, и решение этой мнимой проблемы, которая может стать тяжким препятствием в развитии человеческого познания, является одним из важнейших достижений, к которым приводят независимо друг от друга, но в полном согласии между собой онтология Гартмана и каузально-аналитическое исследование живых систем: одностороннее проникновение слоев или уровней интеграции приводит к тому, что к ним неприменима форма мышления, основанная на взаимно исключающих противоположностях (die Denkform des kontradiktorischen Gegensatzes). В никогда не есть не-А, а всегда А + В, С есть А + В + С и так далее. Но хотя в действительности неуместно подчинять слои реального мира дизъюнктивным понятиям, такие понятия угнездились бесчисленными парами в нашем мышлении, в нашем научном и обиходном языке: природа и дух, тело и душа, животное и человек, естество и воспитание*.
Когда мы в нашем модельном примере (см. с. 271) интегрируем в единую систему две системы (а) и (Ь), т. е. составляем цепь с индукционной катушкой и конденсатором, то это новое единство все еще состоит из обеих подсистем, являющихся его частями, но обладает свойствами, никоим образом даже в зачаточном виде не заметными в этих подсистемах.
Казалось бы, должно быть столь же понятно, что аналогичные новые фульгурации, буквально на каждом шагу происходившие в ходе эволюции, при всей своей новизне оставляли в силе старые системные свойства.
Но есть представители философской антропологии, по-видимому, никак не способные это понять и предающиеся бесконечным бесплодным дискуссиям о том, отличается ли человек от "животного" "по своему существу" или только "степенью". Они не знают или не понимают, что любое вновь возникающее системное свойство, подобно колебательному характеру тока в нашей модели, безусловно, означает не "постепенное", а принципиальное изменение. Теплокровное животное со своей новой управляющей цепью, поддерживающей постоянство температуры, тем самым принципиально отлично от своих предков с переменной температурой, птичье крыло принципиально отлично от конечности пресмыкающегося, из которой оно возникло, и в точности в этом, и ни в каком ином смысле человек принципиально отличен от других антропоидов. Мой учитель Оскар Гейнрот имел обыкновение дружелюбно и терпеливо прерывать каждого, кто в его присутствии говорил в дизъюнктивных терминах о "человеке" и "животном", спрашивая: "Простите, когда вы говорите о животном, имеете ли вы в виду амебу или шимпанзе?"
5. РЕЗЮМЕ ДВУХ ПРЕДЫДУЩИХ ГЛАВ
Я хотел бы выделить из двух предыдущих глав, посвященных возникновению новых системных свойств и учению о слоях Николая Гартмана, три факта, имеющих значение для главной темы этой книги — сравнительного исследования строения и филогенеза когнитивных механизмов. Как и все жизненные процессы, процессы приобретения и накопления информации, способствующей сохранению вида, многослойны и сложно сплетены между собой. При их рассмотрении мы будем все время встречаться со следующими тремя фактами: Во-первых. Простые и простейшие системы вполне способны функционировать самостоятельно, так же как простейшие организмы жизнеспособны и всегда были таковы, потому что иначе из них никак не могли бы возникнуть более высокоорганизованные потомки.
Во-вторых. Новая и сложная функция часто, если не всегда, возникает посредством интеграции нескольких уже имевшихся более простых функций, которые и в отдельности, независимо от этой позднейшей интеграции, способны были к функционированию, а затем никоим образом не исчезают и не теряют своей важности, функционируя в качестве необходимых составных частей нового единства.
В-третьих. Совершенно напрасно искать в отдельных, независимо функционирующих подсистемах или в низших организмах те системные свойства, которые возникают лишь на более высоком уровне интеграции.
Глава 4 ПРОЦЕССЫ ПРИОБРЕТЕНИЯ ТЕКУЩЕЙ ИНФОРМАЦИИ
1. ОГРАНИЧЕНИЯ ФУНКЦИИ ГЕНОМА
Несмотря на почти беспредельную емкость генома, его механизм проб и ошибок сам по себе не смог бы поддерживать живые системы в состоянии непрерывной приспособленности, обеспечивающем их выживание. А именно, когнитивный механизм генома не в состоянии справиться с быстры ми изменениями окружающей среды. В самом деле, он ничего не может "узнать" об успехе какого-либо из своих экспериментов, прежде чем не пройдет свой жизненный путь по меньшей мере одно поколение. Поэтому геном со своими процессами может вырабатывать приспособления лишь к таким условиям окружающей среды, которые сохраняются с достаточным статистическим постоянством в течение длительных периодов времени. На языке современной кибернетики можно сказать, что продолжительность поколения есть "время запаздывания", которое должно пройти, прежде чем когнитивный механизм генома начнет реагировать на внешнее влияние.
Однако, как было отмечено уже в "Пролегоменах" и подробнее рассмотрено в разделе 3 главы 1 (с. 266), существует множество хорошо приспособленных механизмов, принимающих и оценивающих инфор- ·
мацию, но не накапливающих ее. Как уже было сказано выше, их своеобразие часто упускается из виду, поскольку функциональные аналогии между простейшей и самой первичной формой приобретения знания, какую представляет геном, и высшими формами человеческого стремления к познанию, воплощенными в культуре, слишком легко заставляют забывать, что между этими процессами, происходящими на разных уровнях органического бытия, располагается целый слой необходимых когнитивных функций, сообщающих знание о состоянии среды в данный момент и составляющих тем самым основу для всех более высоких видов опыта и обучения. К процессам этого рода мы и должны теперь обратиться. Они существуют у всех живых организмов, в том числе у бактерий и растений.
2. РЕГУЛИРУЮЩИЙ КОНТУР, ИЛИ ГОМЕОСТАЗ
Все физиологические механизмы, получающие кратковременную информацию и тем самым преодолевающие время запаздывания генома, обладают такой способностью благодаря структурам, соответствующая конструкция которых выработана геномом с помощью его метода проб и ошибок. Здесь заключена проблема: даже простейшая форма приобретения текущей информации, а именно регулирующий контур, или гомеостаз, связана со структурой и тем самым с результатами метода проб и ошибок, свойственного геному; а поскольку, с другой стороны, жизнь как таковая, вряд ли мыслима без гомеостаза, то возникает вопрос, отличающийся от известного вопроса о курице и яйце лишь тем, что он имеет смысл.
Мы уже видели на с. 272, что такое регулирующий контур, или цикл с отрицательной обратной связью. В мире организмов существует бесчисленное множество регулирующих контуров всевозможных степеней сложности, начиная от простейших механизмов, поддерживающих некоторое устойчивое состояние на "чисто" химическом уровне, вплоть до высокодифференцированных устройств, в которых сложнейшие функции органов чувств и центральной нервной системы поддерживают, через поведение отдельных индивидов или целых сообществ, определенное "заданное значение", например благоприятную для сохранения вида плотность популяции, как это удалось показать Уинн-Эдвардсу для различных видов животных.
Вообще, когда организм восстанавливает свое внутреннее равновесие после некоторой помехи или сохраняет его вопреки внешним воздействиям, угрожающим этому равновесию, это значит, что он получил и целесообразно оценил информацию о характере и степени соответствующего изменения окружающей его среды. Если, например, животное, находящееся в бедной кислородом среде, учащает свое дыхание или при избытке пищи временно приостанавливает еду, и т. п., это значит, что живая система информирована не только о своей собственной потребности в некоторых веществах, но также о "рыночной конъюнктуре", существующей в данный момент в окружающей среде для этих веществ.
Как и многие другие механизмы, с помощью которых организм получает информацию о текущем положении в окружающем мире,
регулирующий контур действует неограниченно долго без всякого изменения его функции. Иными словами, если отвлечься от нежелательных явлений износа и старения, его запрограммированная геномом структура в течение длительного времени остается неизменной. Информация, для приема и использования которой устроен этот аппарат сразу же оценивается, но не накапливается.
3. СТИМУЛИРУЕМОСТЬ
За исключением некоторых форм гомеостаза все процессы, с помощью которых организм получает и оценивает мгновенную информацию, основаны на его способности отвечать на так называемые стимулы (Reize). Стимул и стимулируемость (Reizbarkeit) определяли весьма различным образом. Вообще под стимулированием понимают такое явление, когда некоторое внешнее воздействие, в смысле причины, запускающей некоторый процесс, но не обязательно с прямым приложением силы. — воздействие, которое и называется "стимулом", вызывает ответ со стороны организма либо в виде некоторого процесса движения (включая любое изменение состояния движения), либо посредством выделения некоторых веществ, т. е. секреции. Когда говорят о стимулируемости, то, во всяком случае для низших организмов, чаще всего имеют в виду "запускаемость" (Auslosbarkeit) движений. Лишь у высших животных разделение труда между нервной и мышечной системами приводит к тому, что эти животные могут принимать и оценивать стимулы, не обязательно отвечая на них немедленным движением.
По-видимому, неизвестно, происходит ли когда-нибудь у самых низших организмов движение без стимуляции. Бактериологи не смогли ответить мне, существуют ли подвижные живые существа, не способные отвечать на стимулы. В принципе можно предполагать, что подвижность, в особенности способность к перемене места, может увеличивать вероятность получения энергии также и в том случае, когда она не сопровождается приобретением информации.
Но у большинства живых существ способность отвечать на стимулы тесно связана со способностью к перемене места, к перемещению. Можно сказать, что первичная и важнейшая функция перемещения состоит в том, что животное избегает опасной ситуации. Возможно, еще более примитивная функция движений тела заключается в том, что организм, стягивая, насколько возможно, свое тело, выставляет против вредных воздействий внешнего мира как можно меньшую поверхность, покрытую сложенной в складки, утолщенной кожей. С этим видом реакции избегания, характерным для многих прикрепленных или медленно движущихся организмов, часто соединяется выделение секретов, служащих для защиты поверхности. Так ведут себя одноклеточные, кишечнополостные (Coelenterata) и другие беспозвоночные, например улитки.
На стимулируемости основаны как те процессы, в которых получается и оценивается лишь текущая информация, без ее накопления (как это было уже описано для некоторых контуров регулирования), так и все те,
которые происходят в центральной нервной системе и составляют основу самых высоких функций обучения и памяти. Об этих функциях пойдет речь значительно позже.
4.АМЕБОИДНАЯ РЕАКЦИЯ
Примечательным образом самая обычная и простейшая реакция на стимулы в мире организмов, движение, может быть направлена в любую сторону трехмерного пространства. Амебоидная клетка, состоящая лишь из "голой" протоплазмы, движется таким образом, что в некотором месте ее наружный слой, эктоплазма, становится тоньше. Затем выпячивается нечто вроде разорванного мешка, которое при дальнейшем, локализованном утончении внешней оболочки вырастает в ложноножку, так называемый псевдоподий. Далее содержимое клетки, следуя направлению наименьшего сопротивления, вливается в псевдоподий, основание которого все более наполняется и утолщается, и таким образом вся клетка постепенно движется в соответствующем направлении. Рост псевдоподиев сопровождается утолщением и стягиванием эктоплазмы на той стороне ползущей амебоидной клетки, которая в процессе движения оказывается сзади.
Процесс образования ложноножек и продвижения в их направлении прежде объясняли, предполагая, что их главной причиной являются изменения поверхностного натяжения. И в самом деле, на искусственных, неживых моделях можно весьма изящно имитировать весь этот процесс, меняя поверхностное натяжение шаровидной капли. Но после длительного наблюдения амеб в достаточно естественной среде я пришел к убеждению, что такое объяснение слишком просто. Как я уже давно утверждал, можно непосредственно наблюдать, как плазма амебы непрерывно переходит из золеобразного состояния в гелеобразное и обратно, оставаясь жидкой внутри клетки в основании ложноножки, где она протекает вперед, и снова застывая, как текущая лава, при соприкосновении с внешним миром, т. е. с водой или дном сосуда. То, что при поверхностном рассмотрении кажется уменьшением поверхностного натяжения и что в самом деле производит такое же механическое действие, — это частичное растворение гелеобразной эктоплазмы, начинающееся изнутри в том месте, где должен образоваться псевдоподий. Это представление, основанное на простых наблюдениях, было впоследствии полностью подтверждено исследованиями Л. фон Гейльбрунна. Когда поверхность подвергается вредному раздражению, отчего она как будто болезненно стягивается и начинает уползать от раздражителя, это происходит вследствие перехода в гелеобразное состояние жидких до этого частей плазмы, непосредственно примыкающих к раздраженному месту. Стягивание вызывается тем, что переход в гелеобразное состояние сопровождается некоторым уменьшением объема протоплазмы, а это производит такое же механическое действие, как увеличение поверхностного натяжения.
Но течение внутренней плазмы, конечно, не может быть обусловлено лишь перепадом давления, вызванным только что описанными процес-
сами. В самом деле, течения протоплазмы известны также и в растительных клетках, заключенных в жесткую целлюлозную оболочку, где давление везде одно и то же.
Когда амебу наблюдают в ее естественной жизненной среде, т. е. не на подложке микроскопа, а свободно движущейся в чашке с культурой, где она живет, то обнаруживается поразительное разнообразие и приспособленность ее поведения. Как говорит лучший знаток простейших Г. С. Дженнингс, если бы она была размером с собаку, можно было бы без колебаний приписать ей субъективное переживание. И единственный способ движения, описанный выше, позволяет амебе справляться со всеми ситуациями окружающей среды. Следует иметь в виду, что с помощью одного и того же механизма амеба "боязливо" бежит от вредного воздействия, стремится к благоприятному воздействию, а в оптимальном случае "жадно" обтекает предмет, от которого исходит позитивный стимул, и поглощает его. Амеба бежит и ест с помощью одного и того же механизма движения!
Приспособительная информация, обусловливающая кажущуюся разумность амебы, основывается исключительно на ее способности избирательно реагировать на весьма различные внешние стимулы; при этом, однако, величина реакции может быть столь разной, что наблюдатель может усомниться в тождестве механизма движения. Амеба, медленно переползающая в более благоприятное место обитания под действием какого-нибудь перепада температуры, кислотности и тому подобного, производит совершенно иное впечатление, чем амеба, "бросающаяся" на добычу или тем более намеревающаяся с кажущейся хитростью захватить несколькими псевдоподиями быстро движущуюся жгутиковую инфузорию. Большое разнообразие поведения и ориентация во всех трех направлениях, достигаемая столь простым способом, придают амебе вид удивительно "разумного" существа. Между тем эти свойства, если можно так выразиться, вовсе не являются "заслугой" амебы, поскольку они происходят от особенностей, присущих протоплазме, как таковой, и тем самым живому организму, состоящему из одной протоплазмы.
Когда же эта Протеева способность создавать в любом месте функциональный передний и задний конец тела приносится в жертву твердой структуре, прежде всего продольно вытянутой обтекаемой форме всех быстро плавающих организмов, тогда возникает совершенно новая проблема: как целесообразно управлять этим быстрым, но жестким кораблем, перемещая его по всем трем направлениям пространства. Среди многоклеточных лишь немногие радиально-симметричные организмы, например морские звезды, способны передвигаться в любую сторону, хотя только в двух измерениях. И если осьминог, это удивительное сказочное существо, при поверхностном наблюдении кажется свободным от ограничений, налагаемых твердыми структурами всех высших организмов, если он в самом деле обладает, подобно амебе, полной свободой перемещения в любом направлении пространства, то он обязан этим не отсутствию структур, но их многообразию и совершенному владению ими.
5.КИНЕЗИС
Теперь нам предстоит рассмотреть ряд механизмов управления, позволяющих животным со структурно закрепленными передним и задним концами целесообразно использовать для сохранения вида свою способность к передвижению, отыскивая места, где более вероятно приобретение энергии и менее вероятна ее потеря. Может показаться удивительным, что это может быть достигнуто без воздействия на направление движения. И все же это возможно. Организм, движущийся в случайном направлении, ускоряющий при этом свое движение, как только окружающие условия становятся неблагоприятными, и замедляющий движение, когда они благоприятны, достигает требуемого эффекта таким чисто количественным воздействием на процесс движения. В виде иллюстрации можно представить себе (нежелательный, впрочем) процесс, когда число автомобилей возрастает в тех местах улицы, где условия замедляют движение. Если бы на их месте были простейшие животные вблизи разлагающейся частицы растения, то их поведение было бы целесообразным. Этот простейший вид поведения, ведущий к возможно более длительному пребыванию в возможно более благоприятных условиях среды, Френкель и Ганн назвали кинезисом (что означает движение).
У многих организмов этот процесс становится еще более эффективным оттого, что животные, движущиеся не прямолинейно, а более или менее зигзагообразно, увеличивают угол между случайно распределенными направлениями движения, как только попадают в благоприятные условия. Как легко понять, таким образом они значительно выгоднее используют пригодную для эксплуатации область. Такой процесс, под названием клино-кинезис, наблюдается большею частью в комбинации с простым кинезисом у самых разнообразных организмов, и отнюдь не только у простейших и беспозвоночных, но — в самом чистом виде — у многих равноногих ракообразных*, принадлежащих, как известно, к высшим ракам. Такие же в формальном и функциональном отношении, но основанные на гораздо более сложных сенсорных и нервных процессах способы поведения имеются у млекопитающих: достаточно представить себе пасущихся жвачных или людей, ищущих грибы.
Кинезис в собственном смысле замечателен простотой своего механизма. Здесь достаточен единственный рецептор, чисто количественным образом действующий на единственный способ движения. Это, насколько я могу понять, есть простейший процесс, с помощью которого организм, способный не амебоидно передвигаться во всех направлениях, может получать и оценивать текущую пространственно ориентирующую информацию. То, что организм узнает о внешнем мире, можно выразить простыми словами: "Здесь лучше" или "Здесь не так хорошо". Следствия, которые он выводит из этого "знания", столь же просты: "Здесь мы еще побудем" или "Отсюда надо поскорее уйти". При этом животное ничего не узнаёт о направлении перепада*, в котором внешняя среда становится лучше или хуже
6. ФОБИЧЕСКАЯ РЕАКЦИЯ
Есть ряд низших организмов, реагирующих стереотипной реакцией обращения, когда перемена места приводит их к перепаду стимула, означающему быстрое ухудшение условий среды. В таком случае организм узнает нечто и о направлении, в котором находится подлежащее избеганию. Но если, наоборот, животное встречается с изменением, означающим улучшение жизненных условий, то реакция отсутствует, если только не действует кинезис, как это бывает у многих простейших. И лишь когда глупое существо выходит из благоприятной области на другую сторону, т. е. в неблагоприятную среду, оно отвечает на это реакцией избегания; по выражению Отто Кёлера, оно ведет себя точно так же, как поступает человек, кладущий в карман прибавку к заработной плате, не проронив ни слова, но устраивающий большой скандал при любом сокращении его дохода.
Как показал Г. С. Дженнингс и как подчеркивает Альфред Кюн в своей классической книге "Ориентация животных в пространстве", величина угла поворота животного от неблагоприятного направления не зависит от направления воспринятого стимула. Например, туфелька (Paramaecium) при своей реакции поворота ведет себя следующим образом. Сначала реснички на всей ее поверхности начинают работать в обратном направлении, так что она проплывает некоторое расстояние назад по тому же пути, по которому двигалась вначале. Затем реснички на одной стороне тела, а также в окрестности рта, подгоняющие ко рту пищу, начинают снова работать "вперед". Вследствие этого туфелька сначала не движется ни вперед, ни назад, а описывает передним концом окружность, причем продольная ось ее тела движется по боковой поверхности конуса. После некоторого промежутка времени, зависящего от силы стимула, но не от его направления, реснички начинают работать, как вначале, и туфелька плывет в направлении, которое занимала в этот момент ее продольная ось. Может случиться, что при возобновлении движения вперед ее тело повернулось по конической поверхности как раз на 360 градусов; тогда она опять плывет навстречу возрастающему стимулу. Может случиться и так, что новое направление оказывается еще менее благоприятным, чем прежнее, и ведет к еще более крутому нарастанию пугающего стимула. В обоих случаях животное повторяет свою реакцию. Этот способ поведения, названный Альфредом Кюном фобической реакцией, в отношении объема доставляемой животному информации значительно превосходит кинезис (но никоим образом не амебоидную реакцию). Животное не только узнаёт при этом, что некоторое место неблагоприятно, но также в каком направлении условия еще менее благоприятны, хотя и не получает сведений о том, в каком направлении нежелательные условия хуже всего и тем более в каком направлении надо искать благоприятные условия. Поскольку фобическая реакция не только оказывает количественное воздействие на движение. как это делает кинезис, но вызывает сверх того еще качественно иную реакцию обращения, животное может в течение длительного времени избегать неблагоприятной среды и оставаться в благоприятной,
а не только сокращать свое пребывание в первой и удлинять во второй, как это позволяет делать кинезис.
Информация о том, какова благоприятная и неблагоприятная среда, при фобической реакции доставляется организму, так же как при амебоидной реакции и кинезисе, на той стадии общего процесса, на которой стимулы принимаются и в определенном смысле фильтруются. Проблемы, связанные с этой функцией, я рассмотрю в восьмом разделе этой главы'.
7. ТОПИЧЕСКАЯ РЕАКЦИЯ, ИЛИ ТАКСИС
На гораздо более высоком уровне, как в отношении количества получаемой текущей информации, так и в смысле сложности участвующих процессов, стоит тип реакций ориентации, которые мы назовем вместе с Альфредом Кюном топическими реакциями, или таксисами. После опубликования его уже упомянутого труда было предпринято много важных исследований о пространственной ориентации животных. Для понимания лежащих в ее основе физиологических процессов особенное значение имели те из них, которые основываются на кибернетической точке зрения, на учении о регулирующем контуре, такие, как работы Миттельштедта, Яндера и других. Рамки этой книги не позволяют дать сводное изложение их результатов.
Простейшая топическая реакция, в терминологии Кюна — тропо-таксис, состоит в том, что организм вращается до тех пор, пока между двумя симметрично расположенными рецепторами устанавливается равновесие стимулов. Плоский червь, реагирующий "положительным тропо-таксисом" на течения, приносящие ему запах пищи, вращается до тех пор, пока поток воды с обеих сторон его головного конца достигает равной силы, а затем ползет против течения. Если искусственно устроить такую ситуацию, симметрично направив на голову червя два водяных потока из раздвоенной трубки, то червь проползает между ними по результирующему направлению. От этого простого механизма, почти соответствующего предложенной Лёбом абстракции "тропизма", все мыслимые переходы ведут к нервным организациям со сложнейшими обратными связями; хорошим примером этого является аппарат, ' *Не следует думать, что фобическая реакция является единственным механизмом ориентации, свойственным парамеции. При более слабом перепаде, а также в случае, когда животное входит в область более сильного перепада под большим углом к нему, оно вполне способно — как показала Вальтрауд Розе — в целесообразной степени изменить свой курс и избежать неблагоприятной среды, не прибегая к фобической реакции. В естественных условиях фобическая реакция наблюдается лишь изредка. Как показал Отто Кёлер, туфелька способна также различать стимулы, затрагивающие ее переднюю и заднюю части. Лишь на первые она отвечает фобически, на последние же — вполне целесообразным ускорением движения вперед. Эта способность различения отказывает при сверхсильных стимулах. Если внезапно поднести к заднему концу туфельки очень горячую иглу, она совершает точно такой же прыжок назад, как при соответствующем раздражении переднего конца. Такой способ поведения, названный Отто Кёлером реакцией испуга, может оказаться для животного гибельным; впрочем, это происходит лишь в условиях, вряд ли встречающихся в его нормальной свободной жизни.
с помощью которого богомол (Mantis) направляет свою атаку на добычу, как это сумел показать Г. Миттельштедт в итоге многолетних исследований.
Все эти топические реакции, от простейших до самых сложных. имеют ту общую черту, что животное сразу же, без проб и ошибок. выбирает пространственное направление, благоприятное для сохранения вида. Иными словами, величина угла, на который поворачивается животное, непосредственно зависит от угла между воспринятым стимулом и продольной осью животного. "Отмеренный" поворот характерен для всех топических реакций.
В то время как фобическая реакция дает организму информацию только о направлении, в котором он не должен двигаться, ничего не говоря ему о бесчисленных других направлениях пространства, которые он мог бы избрать, топическая реакция непосредственно информирует животное, какое из всех этих возможных направлений наиболее благоприятно. Таким образом, в отношении получаемой информации таксис многократно превосходит и фобическую реакцию, и кинезис, но, подчеркнем еще раз, не реакцию псевдоподий амебоидной клетки.
8. ВРОЖДЕННЫЙ МЕХАНИЗМ ЗАПУСКА
В разделах, посвященных движению амебы, кинезису и фобической реакции, я упомянул уже наряду с функцией приобретения информации, выполняемой соответствующим процессом движения, также функцию физиологического механизма, запускающего этот процесс. Организм нуждается не только в структурах, обеспечивающих моторное* осуществление некоторой формы движения, способствующей сохранению вида; ему нужен также аппарат для приема стимулов, говорящих ему, в какой момент и при каких обстоятельствах соответствующая форма поведения имеет шанс выполнить свое назначение.
Когда в физиологии нервной системы был открыт важный принцип рефлекторной дуги, казалось естественным охватить понятием рефлекса все процессы, запускающие движение; а когда И. П. Павлов объяснил не менее важный процесс возникновения условных реакций, казалось естественным истолковать все врожденные целесообразные реакции, т. е. реакции, происходящие без предварительного обучения и полезные для сохранения вида, как "безусловные рефлексы". Само по себе такое представление не ошибочно, но оно заслоняет подлинную проблему. Для животного с центральной нервной системой весьма вероятно, что аппарат, принимающий стимулы, так называемый рецептор, связан в одну систему с эффектором, т. е. нервным устройством, осуществляющим целесообразный моторный ответ, посредством нервного пути, хорошо подходящего под общее понятие рефлекторной дуги. В целом ряде случаев точно изучено, как проходит этот путь и из каких нервных элементов он состоит.
Но наша проблема заключается не в самом процессе рефлекса, а в некотором смысле в том, что ему предшествует, в его рецепторном начале. Мы должны спросить себя: как получается, что организм в точности "знает", какая именно реакция должна последовать за данным
стимулом, чтобы осуществилась функция, полезная для сохранения вида? Каким образом получается, например, что амеба обволакивает и поглощает не все мелкие частицы, но — за редкими исключениями — только те, которые могут служить ей пищей? Откуда знает маленькое существо, пробивающееся через жизнь с помощью кинезиса, когда и где оно должно плыть быстро или медленно?
Следует предположить, что каждому такому моторному ответу предшествует работа механизма, фильтрующего стимулы, т. е. позволяющего действовать лишь тем из них, которые с достаточной статистической достоверностью характеризуют внешнюю ситуацию, где запускаемый способ движения .может оказаться целесообразным. Этот рецепторный аппарат можно сравнить с замком, который отпирается лишь вполне определенным ключом. Поэтому употребляется также выражение ключевой стимул. Физиологический аппарат, фильтрующий стимулы, мы назовем врожденным механизмом запуска, сокращенно — ВМЗ.
У одноклеточных и низших многоклеточных с их не слишком богатым запасом различных форм движения, по существу ограничивающимся поиском добычи и полового партнера, а также избеганием опасных ситуаций, к избирательности ВМЗ предъявляются не слишком высокие требования. И все же амеба избирательно реагирует на целый ряд стимулирующих ситуаций, хотя ее формы поведения различаются лишь количественно. По сравнению с нею заключенные в жесткую структуру жгутиковые инфузории, к которым принадлежит парамеция, кажутся далеко не столь пластичными. Это животное отыскивает с помощью своих фобических и топических реакций среду с требуемыми свойствами, и прежде всего с определенной концентрацией Н-ионов. Чаще всего встречающаяся в природе кислота есть CO2*, и ее повышенная концентрация обнаруживается в водах, где находят парамеций, чаще всего поблизости от гниющих остатков растительных веществ; кислоту выделяют скопления бактерий, питающихся этими остатками. Связь эта столь надежна, а присутствие других, особенно ядовитых, кислот столь редко, что парамеция отлично обходится очень простой информацией, которую можно выразить словами: где имеется определенная концентрация кислоты, там собираются съедобные бактерии. Разумеется, программа вида не предусматривает экспериментирующего физиолога, который вводит в жизненное пространство парамеции каплю ядовитой щавелевой кислоты.
У высших животных с хорошо развитыми центральной нервной системой и органами чувств, а также с богатым запасом качественно различных форм поведения, к избирательности врожденных механизмов запуска предъявляются более высокие требования, особенно в тех случаях, когда различные комбинации стимулов, воспринимаемые одним и тем же органом, должны вызывать разные ответы. Когда, как, например, в случае самки сверчка, некоторый орган воспринимает лишь единственный вид раздражения, вызывающий единственное ответное поведение, эта проблема не возникает. Как показал Реген, самка сверчка не слышит ничего, кроме призыва самца сверчка. Напротив, мальки
большинства видов цихлид оптически реагируют как на образ их матери, за которой они следуют, так и на хищную рыбу того же размера, от которой они спасаются бегством в укрытия. Любой из этих двух способов поведения. Примененный к неправильному объекту, означал бы несомненную гибель.
В указанном случае самки сверчка орган слуха теоретически мог бы быть прямо соединен с исполнительным моторным аппаратом. Но в случае рыб между рецептором и эффектором должен быть фильтрующий аппарат, способный различать два вида ключевых стимулов. Сам он может быть расположен только в нервной системе, т. е. между воспринимающим и исполняющим органами.
О том, каким образом ВМЗ осуществляет свою физиологическую фильтрацию, мы знаем очень мало, хотя исследования Леттвина с сотрудниками, а также Экгарда Бутенандта на сетчатке лягушки бросают яркий свет на возможности такой сортировки. В последнее время Швартцкопф и его ученики показали на кузнечиках, что цепь ганглий, которую должны проходить ключевые стимулы, в самом деле выполняет фильтрацию, и установили, как это происходит.
Когда мы видим в естественных условиях, с какой уверенностью и целесообразностью ВМЗ сообщает организму, какие именно способы поведения способствуют в данных обстоятельствах сохранению вида, возникает тенденция к переоценке количества информации, заключенного в таком сообщении. Когда мы видим, как "разумно" ведут себя парамеции вблизи толчеи кормящихся бактерий, или как только что вылупившийся индюшонок при виде пролетающей хищной птицы забивается в ближайшее укрытие, или как молодая пустельга* при первом столкновении с водой купается в ней и затем чистит свои перья, как будто она уже делала это тысячу раз, то мы узнаем почти с разочарованием, что примитивные инфузории ориентируются только по концентрации кислоты, что индюшонок точно так же прячется от большой мухи, ползающей по белому потолку, и что гладкая мраморная плита вызывает у молодой пустельги те же движения, что вода.
Врожденная информация механизма запуска закодирована столь просто, как это только возможно при условии, что в биологически неадекватных ситуациях его действие должно быть маловероятным. Классическим примером простой, но вполне достаточной для животного в естественных условиях информации служит ВМЗ, вызывающий реакцию укуса у обыкновенного клеща (Ixodes rhicinus). Как показал Якоб фон Юкскюль, клещ кусает все, что имеет температуру в 37° С и пахнет масляной кислотой. Насколько проста эта характеристика естественного хозяина клеща, млекопитающего, настолько же невероятно, чтобы эту реакцию мог вызвать какой-нибудь другой встречающийся в лесу предмет.
Одно из самых основательных и точных исследований ВМЗ было проведено на мальках цихлид супругами Кюнцер. Хороший обзор современного состояния этой проблемы содержится в работе В. Шлейдта (1964).
9. СВОЙСТВЕННОЕ ВИДУ ИМПУЛЬСИВНОЕ ПОВЕДЕНИЕ В СМЫСЛЕ ОСКАРА ГЕЙНРОТА*
Врожденный механизм запуска играет особую роль, когда он приводит в действие так называемое инстинктивное движение. У организмов, которым жесткий, расчлененный скелет оставляет лишь вполне определенные степени свободы, т. е. прежде всего у членистоногих и позвоночных, всегда есть свойственные виду двигательные координации, запрограммированные в геноме как одно целое и готовые к выполнению. По-немецки они называются наследственными координациями (Erbko ordinationen), или инстинктивными движениями (Instinktbewegungen), по-английски "закрепленными шаблонами движения" ("fixed motor patterns"). Физиологически они характерны тем, что их очень жесткая последовательность движений порождается не сцеплением рефлексов, как естественно было бы предположить, а процессами, происходящими в самой нервной системе без участия рецепторов. Эрих фон Гольст, Пауль Вейс и другие посвятили подробные исследования физиологии центрально координированных форм движения. Как показали недавно Э. Тауб и А. Дж. Берман, даже у приматов большая часть их часто высокодифференцированных двигательных координации функционирует независимо от какого-либо управления внешними и внутренними рецепторами. При врожденных координациях афферентный* контроль играет роль лишь в общей пространственной ориентации, но несуществен для самого возникновения последовательности движений. Это можно уяснить себе даже без вивисекционных опытов дезафференции (т. е- выключения всех чувствительных нервов) по часто происходящим холостым движениям, в которых врожденная координация выполняется в целости без присутствия нормально вызывающего ее объекта. Так, например, ткачик, Quelia, может выполнять все сложное движение, служащее для закрепления на ветке соломинки при постройке гнезда, даже при отсутствии соломинки или какого-либо подобного предмета. Это поведение выглядит так, как будто птица "галлюцинирует" отсутствующий предмет.
Выполнение наследственной координации, рассматриваемое само по себе, не есть когнитивный процесс. Содержащееся в ней готовое к употреблению моторное умение находится в распоряжении животного как хорошо сконструированное орудие, и чем более специализировано это орудие, тем уже область его применения. Есть врожденные координации общего назначения, как, например, координации перемены места, грызения, чесания, долбления, и т. д., и есть другие, в высшей степени специализированные для определенной функции, как, например, уже упомянутое связывающее движение ткачика или многие формы поведения при токовании и оплодотворении.
Именно в этих врожденных координациях, дифференцированных для вполне определенных функций, наиболее отчетливо проявляется их точно приспособленная жесткость, их полная независимость от какого-либо обучения. Даже опытный этолог снова и снова удивляется, видя, как только что выращенное молодое животное, о котором достоверно известно, что оно не могло получить информацию из собственного опыта,
впервые демонстрирует такую последовательность поведения во всей ее целесообразности и совершенстве. Оскар Гейнрот описывает, как выращенный из яйца и едва научившийся летать ястреб поймал в воздухе фазана, пытавшегося перелететь со стола на подоконник, и прежде чем смог вмешаться его воспитатель, уселся с уже убитой добычей на шкаф. Гейнрот прибавляет: "Это первое профессиональное действие ястреба произвело на нас неизгладимое впечатление". В действительности соединение моторного умения и точного "знания" ситуации, в которой это умение должно быть применено, предполагает огромную массу врожденной информации.
Только что описанная форма поведения, состоящая из срабатывания некоторого ВМЗ и запущенного им действия врожденной координации, образует функциональное целое, чрезвычайно часто встречающееся в царстве животных. Оскар Гейнрот назвал его "свойственным виду импульсивным поведением". Это понятие оказалось чрезвычайно плодотворным, и лишь много времени спустя дальнейший анализ этого единства показал, что две его компоненты могут интегрироваться в функциональное целое также и другим способом.
У высших животных свойственное виду импульсивное поведение представляет собой прототип когнитивного процесса, который, как уже говорилось в главе 1 (см. с. 267), является не приспособлением, а функцией уже приспособленного механизма. При рождении организму задается информация о биологически "правильных" ситуациях и о средствах, позволяющих ему справляться с такими ситуациями. Процесс, доставляющий текущую информацию, говорит животному лишь одно: "Hic Rhodus, hic salta"'*, теперь наступил момент применить вот этот особенный способ поведения!
Свойственное виду импульсивное поведение является типичным примером линейной цепи актов поведения, пригодной к функционированию уже в этой простой форме; но при интеграции с другими такими цепями, каждая из которых столь же проста, возникает "фульгурация" поистине эпохальных новых функций. Простая цепь описанного выше типа встречается, собственно, лишь у тех живых существ, у которых, как, например, у пауков-скакунов и многих насекомых, такое поведение осуществляется в жизни индивида лишь единственный раз.
Но у высших животных к этому прибавляется еще по крайней мере один дальнейший тип поведения — поиск запускающей ситуации стимулирования, который мы назовем, вместе с Уоллесом Крейгом, аппетентным поведением. Можно предположить, что чисто линейная цепь поведения, дополненная этим предварительным членом, уже имеет значение для сохранения вида. Впрочем, я не могу подтвердить это каким-либо конкретным примером. Почти во всех случаях, когда наблюдается аппетентное поведение, обнаруживается также обратное влияние успеха на предшествующее поведение. Но тем самым возникает тот же круговой процесс, на котором основывается обучение в собственном смысле этого слова, т. е. обучение посредством успеха, о чем будет речь в главе 6.
1* "Здесь Родос, здесь прыгай" (лат.), пословица, взятая из басни Эзопа. — Примеч. пер.
10. ДРУГИЕ СИСТЕМЫ, ПОСТРОЕННЫЕ ИЗ ВРОЖДЕННЫХ МЕХАНИЗМОВ ЗАПУСКА И ИНСТИНКТИВНЫХ ДВИЖЕНИЙ
Как я уже говорил, мой учитель Оскар Гейнрот и я сам в течение долгого времени рассматривали "свойственное виду импульсивное поведение" как простейший и важнейший составной элемент всего животного и человеческого инстинктивного поведения. Понимание, что оно состоит из двух физиологически различных частей, непосредственно следовало из открытия Эриха фон Гольста, убедительно показавшего, что врожденная координация не состоит, как до тех пор считалось само собою разумеющимся, из цепей безусловных рефлексов. Как показал Гольст, координация движений не только выполняется в точной последовательности без помощи рефлексов, но может также начаться без всякого внешнего стимула. Лини, у которых были перерезаны задние корни всех спинномозговых нервов, выполняли вполне нормальные плавательные движения; нервная система дождевого червя, полностью отделенная от остального тела и подвешенная в физиологическом растворе, неуклонно посылала последовательность нервных импульсов, которая побудила бы мышечную систему червя, если бы она была, выполнять координированные движения ползания. Таким образом, движение вызывается стимуляцией и координацией, производимыми в самой центральной нервной системе. Как выразился Эрих фон Гольст, "мантия рефлексов" служит лишь для того, чтобы целесообразно приспособить стимулируемые изнутри движения к обстоятельствам места и времени окружающего мира.
Наследственная координация образует неизменный остов поведения, структура которого содержит исключительно филогенетически полученную информацию. Она приводится в действие лишь многочисленными служащими ей механизмами, принимающими текущую информацию, которые в адекватной ситуации запускают эту координацию и направляют ее во времени и пространстве. Это открытие я рассматриваю как момент рождения этологии, поскольку оно доставило Архимедову точку опоры, на которой основано наше аналитическое исследование.
Описанные здесь физиологические открытия бросают новый свет на процессы, образующие совместно с врожденной координацией уже рассмотренные функциональные устройства. До тех пор, пока свойственное виду импульсивное поведение считали цепью безусловных рефлексов, процесс его запуска казался первым членом этой цепи, таким же "рефлексом", как другие. Физиологически он казался не отличающимся от этих других, а потому не заслуживающим особого внимания. Но когда оказалось, что эндогенная стимуляция каждой такой последовательности движений непрерывно происходит сама по себе и должна постоянно тормозиться особыми контролирующими механизмами, т. е. когда стало ясно, что запуск инстинктивного движения, по существу, означает лишь снятие торможения его спонтанности, тем самым возник вопрос, какой особый физиологический механизм осуществляет такое снятие
торможения.
У многих низших животных важнейшая функция высших инстанций нервной системы состоит именно в том, чтобы осуществлять постоянное
торможение различных свойственных организму эндогенных автоматических движений, а в надлежащий момент, на основании поступающей извне мгновенной информации, "снимать с них узду". Дождевой червь, лишенный своего "мозга", т. е. его верхнеглоточной ганглии, непрерывно ползает и не может остановиться. Подобным же образом оперированный краб не может перестать есть, пока имеется что-нибудь съедобное, и т. п.
Открытие эндогенной стимуляции центрально координированных форм движения бросило новый свет не только на процесс их высвобождения, но и на ряд других, иначе устроенных и в высшей степени важных явлений. Из наблюдений Гейнрота, Лисмана и моих собственных давно уже было известно, что при длительном неупотреблении некоторого инстинктивного движения порог вызывающего его стимула не остается постоянным, а все более снижается. Вследствие этого соответствующая форма поведения запускается все легче, начинает срабатывать в ответ на неадекватные стимулы, на "замещающие объекты", и в экстремальном случае оно происходит наконец без всякого заметного стимула — •как мы говорим, "вхолостую" ("auf Leerlauf). Это явление снижения порога я изучил и описал, когда еще твердо верил, что инстинктивные движения представляют собой цепи рефлексов. Но уже в то время я добросовестно уделил особое внимание тем явлениям, которые нельзя объяснить теорией рефлекторных цепей. И как раз эти явления получили яркое объяснение благодаря открытиям Гольста. Впрочем, снижение порога запускающего стимула — не единственное следствие длительного отсутствия ситуации, адекватной для определенного инстинктивного движения. Длительное лишение условий выполнения некоторой врожденной координации большею частью приводит организм как целое в состояние беспокойства, побуждая его активно искать ключевые стимулы. Это уже рассмотренное в конце предыдущего раздела явление мы называем, следуя Уоллесу Крейгу, аппетентным поведением (appetitive behavior).
То, что "подгоняет"* животное, "импульс", неявно содержащийся в старом термине Гейнрота "импульсивное поведение", таким образом, во многих случаях вовсе не совпадает с одним из "великих" общих побуждений животного и человеческого поведения — таких, как голод, жажда или половое влечение, — физиологические причины которых сравнительно легко обнаруживаются и состоят в нехватке важных веществ либо в переполнении пустотелых органов ("импульс детумесценции"*). Более того, каждое мельчайшее специальное инстинктивное движение представляет собой автономный стимул поведения, если оно в течение некоторого времени не "отреагируется" ("abreagiert") — если воспользоваться этим не очень изящным выражением Зигмунда Фрейда. Это играет важную роль в образовании условных реакций, о чем будет сказано в главе 6. Как впервые показали Н. Тинберген и Г. П. Берендс, три важнейших подсистемы стимулируемого поведения: аппетентное поведение, действие врожденного механизма запуска и успокаивающее выполнение инстинктивного движения — могут соединяться друг с другом также иным, более сложным образом. Очень часто продолжительная последовательность форм поведения программируется таким образом, что составляющее ее начало аппетентное поведение доставляет
стимулирующую ситуацию, которая запускает не непосредственно инстинктивное движение, составляющее его цель, а вначале лишь другой вид аппетентного поведения. Чеглок* летает в поисках добычи — это аппетентное поведение первого порядка. Он встречает стаю скворцов и, высоко поднявшись над нею, выполняет особый маневр, имеющий целью отрезать от стаи одного определенного скворца, — это аппетентное поведение второго порядка. Лишь в случае, если это поведение приводит к успеху, хищник достигает ситуации, в которой осуществима следующая форма поведения, а именно умерщвление добычи, за которым следуют дальнейшие инстинктивные движения — сначала ее ощипывание, а затем пожирание. Для нашего представления о сущности инстинктивного движения важно то обстоятельство, что многие врожденные координации встроены в такую последовательность не в качестве удовлетворяющей инстинктивное стремление конечной цели, а в некотором смысле в виде промежуточных целей. Их можно поэтому с равным правом истолковать как формы аппетентного поведения, направленные к достижению стимулирующей ситуации, запускающей следующий член последовательности. Такую последовательность аппетенций Тинберген назвал иерархически организованным инстинктом.
Такая система иерархически упорядоченных аппетенций по филогенетически "хорошо продуманной" программе ведет животное от одной стимулирующей ситуации к другой, причем — ив этом суть дела — всегда от более общей, легче находимой к более специальной, которую трудно было бы найти, если бы первый механизм запуска, производящий переключение с аппетенций первого порядка на аппетенцию второго порядка, не указывал к ней путь. Понятие иерархически организованного инстинкта не имеет ничего общего с метафизическими конструкциями: число участвующих в нем механизмов запуска можно определить экспериментально, а участвующие врожденные координации
допускают точное описание.
Цепи аппетенций и механизмов запуска, участвующих в такой системе, могут также образовывать "вилку", когда после достижения стимулирующей ситуации, к которой ведет аппетентное поведение первого порядка, имеется возможность нескольких дальнейших способов поведения, т. е. пробуждается несколько дальнейших аппетенций. Самец колюшки, у которого весной, с возрастанием продолжительности дня, гормональным путем развивается готовность к размножению, начинает сначала перемещаться из глубокой воды в мелкую и продолжает это аппетентное поведение до тех пор, пока не оказывается в заросшем растениями спокойном мелководье. В этой ситуации он становится "территориальным", т. е. завладевает вполне определенной областью, которую более не покидает. При этом меняется его "установка", т. е. готовность к определенным формам поведения, что внешним образом сообщается как собратьям по виду, так и исследователям его поведения изменением его окраски. С этого момента самец колюшки в равной мере готов строить гнездо, сражаться с соперниками и ухаживать за самкой, вести ее к гнезду и там с нею нереститься. Таким образом, стимулирующая ситуация, достигнутая аппетентным поведением первого порядка, составляет предпосылку для одновременного возникновения нескольких
различных аппетенций. Систему инстинктов колюшки глубоко изучил Н. Тинберген, а для случая песочной осы (Ammophila) эту работу проделал Г. П. Берендс.
Подобная иерархически организованная система инстинктов намного "пластичнее", чем простое импульсивное действие в смысле Гейнрота, поскольку каждый из многих последовательно включаемых механизмов запуска получает информацию об условиях окружающей среды в данный момент и приспосабливает к ним поведение. Сверх того многие участвующие в общем процессе и направляющие его в пространстве таксисы также вносят значительный вклад в приобретение информации.
Иерархические системы инстинктов обладают особой приспособительной способностью, экономящей время и энергию; она состоит в том, что в определенных случаях отдельные члены цепи поведения могут опускаться. Когда чеглок в нашем предыдущем примере сталкивается с отдельным скворцом, то он, конечно, не производит маневр отрезания от стаи и переходит прямо к умерщвлению. Как показал Берендс, соответствующие "сокращения" не всегда удается вызвать у песочной осы; во многих случаях насекомое застревает в обязательной иерархической последовательности действий.
Подчеркнем еще раз, что эта поистине удивительная и тем самым производящая впечатление "разумности" приспособительная способность иерархически организованной системы достигается посредством функций, служащих для приема текущей информации. Система приспособлена, но она обеспечивает своей открытой программой богатый набор комбинаций, целесообразный для сохранения вида даже и без адаптивной модификации ее механизма. Тем самым она полностью относится к категории когнитивных процессов, действующих без обучения, — процессов, составляющих предмет этой главы. Независимость их действия от обучения может быть доказана в тех многих случаях, где сложные, иерархически организованные цепи поведения встречаются в жизни индивидуального животного лишь единственный раз, как, например, в столь основательно изученном супругами Пекхемом и Джоселин Крейн поведении спаривания многих пауков, или исследованном и заснятом на кинопленку Эрнстом Ризом поведении личинок рака-отшельника, которые уже в стадии свободно плавающей Glaucothoe при первой же встрече с раковиной улитки выполняют всю высокодифференцированную последовательность форм поведения, с помощью которых взрослый рак-отшельник находит, обследует, чистит и заселяет раковину.
Принципиальная независимость действия иерархически организованного инстинктивного поведения от процессов обучения не исключает того факта, что как раз оно и стало основой, на которой развились механизмы обучения. К высокоразвитым процессам обучения оно находится в том "одностороннем" отношении, которое характерно для явлений, происходящих на различных уровнях интеграции. Вспомним, что было сказано об этом выше в разделе 4 главы 2 (см. с. 274).
Механизмы, получающие текущую информацию, независимы в своем действии от процессов обучения, добавляющихся на высшем уровне, но составляют предпосылку для их возникновения. Без них были бы
невозможны рассматриваемые в следующих двух главах процессы адаптивной, или "телеономной", модификации поведения путем обучения. Прокладывание путей посредством упражнения, ослабление стимулов вследствие привыкания, усиление избирательности механизмов запуска вследствие привычки — короче, все те процессы, о которых пойдет речь в следующей главе, возможны лишь на основе функций, описанных в этой главе. Это в особенности справедливо в отношении когнитивной функции обучения посредством успеха (conditioning by reinforcement), которое будет рассмотрено в главе 6. Возникновение этой высшей и важнейшей формы обучения имело своей предпосылкой наличие вполне пригодных к действию систем, состоявших из аппетентного поведения, врожденного механизма запуска и конечной целевой ситуации. Без наличия этих трех членов никогда не могла бы возникнуть "фульгурация" той обратной связи, посредством которой успех воздействует в обратном направлении на предшествующее поведение и которая составляет сущность условных реакций в узком смысле.
11. РЕЗЮМЕ ГЛАВЫ
Из когнитивных механизмов, рассмотренных в первых трех главах, только механизм генома с его методом проб и ошибок в состоянии не только приобретать информацию, но и накапливать ее. Количество приобретаемой и сохраняемой таким образом информации почти безгранично, но время, необходимое для того, чтобы вновь полученное знание привело к полезным для сохранения вида последствиям, соответствует по меньшей мере жизни поколения. Поэтому живые системы — если они не находятся в практически невозможной неизменной среде — могут сохранять свою приспособленность лишь при участии краткосрочно действующих механизмов, дающих информацию о текущем состоянии окружения и оценивающих полученные данные.
Самый первоначальный, древнейший и вездесущий из этих механизмов — это регулирующий контур, посредством обратной связи делающий внутренние условия организма независимыми от колебания внешних условий и поддерживающий их постоянство (гомеостаз).
У свободно движущихся организмов имеются процессы, позволяющие им целесообразно справляться с пространственными условиями окружающей их среды, такие, как, например, амебоидная реакция, кинезис, фобическая реакция и таксис, общей предпосылкой которых является способность воспринимать стимулы.
На более высоком уровне дифференциации за ними следуют врожденный механизм запуска, наследственная координация, или инстинктивное движение, и более сложные системы, построенные из двух предыдущих.
В противоположность когнитивным функциям генома, а также высшим функциям познания, включающим в себя обучение, все описанные в этой главе механизмы не способны накапливать информацию. Действие их представляет собой не процесс приспособления, а функцию готовых приспособленных структур. Они защищены от любой модификации и должны быть таковыми, поскольку предшествуют любому опыту,
составляя основу всякого возможного опыта. В этом отношении они соответствуют "априорному", как его определил Кант.
Рассмотренные в этой главе механизмы приобретения текущей информации, способные действовать независимо от какого-либо обучения, составляют, в свою очередь, необходимую основу для процессов обучения, развивающихся на высшем уровне интеграции.
Глава 5 ТЕЛЕОНОМНЫЕ МОДИФИКАЦИИ ПОВЕДЕНИЯ
(за исключением обучения посредством вознаграждения — conditioning by reinforcement)
1. ОБЩИЕ СВЕДЕНИЯ ОБ АДАПТИВНОЙ МОДИФИКАЦИИ
Модификацией называется любое изменение свойств организма, вызванное воздействием внешних условий на его индивидуальную жизнь. Для любого живого существа модификация обусловливает, на основе его наследственных задатков, т. е. генотипа, его внешний образ — фенотип. Модификация — это вездесущий процесс. Вряд ли будет преувеличением утверждать, что любое малое различие в условиях окружающей среды, в которой вырастают два генетически одинаковых индивида, влечет за собой некоторое малое различие в их свойствах, т. е. в их фенотипе. Но такие модификации плана строения под влиянием среды вовсе не обязательно суть изменения, полезные в смысле сохранения вида. Напротив, вероятность того, что модификация, вызванная определенным изменением среды, означает приспособление к этому же изменению, не выше вероятности того, что некоторая случайная мутация или рекомбинация генов принесет с собой какое-либо преимущество для сохранения вида. Но если в ответ на некоторое вполне определенное внешнее влияние регулярно происходит модификация, представляющая собой телеономное, т. е. способствующее сохранению вида, приспособление именно к этому влиянию, то можно предположить с вероятностью, близкой к достоверности, что соответствующая специфическая модифицируемость уже является результатом предшествующего отбора.
Если, например, на большой высоте, с уменьшением содержания кислорода при низком давлении воздуха, человеческая кровь обогащается гемоглобином и красными кровяными тельцами; или если собака в холодном климате приобретает более густую шерсть; или если растение, растущее при слабом свете, вытягивается в длину и тем самым доставляет своим листьям лучшее освещение, все эти адаптивные модификации никоим образом не являются следствием только внешнего влияния, которое их вызывает, но также и некоторой встроенной генетической программы, выработанной геномом по методу проб и ошибок и составляющей в каждом отдельном случае уже готовое приспособление к среде. В словесной форме данная растению инструкция звучала бы примерно так: при недостаточном освещении стебель должен вытяги-
ваться до тех пор, пока света не станет достаточно. Следуя Эрнсту Майру, мы назовем этот вид генетической информации открытой программой.
Открытая программа — это когнитивный механизм, способный не только приобретать, но и накапливать информацию о внешней среде, не заключенную в геноме. Иными словами: онтогенетическое применение самой подходящей из способностей, содержащихся в открытой программе, есть процесс приспособления. Таким образом, открытая программа приобретает и хранит информацию; но не следует упускать из виду, что для этой функции ей требуется количество генетической информации не меньшее, а большее, чем в случае замкнутой программы. Это можно пояснить сравнением. Предположим, что человек хочет построить домик из готовых деталей, не требующих никаких предварительных приспособительных изменений, — это пример полностью замкнутой программы. Единственная строительная площадка, на которой можно осуществить такое намерение, — это совершенно плоская поверхность, вроде тех строго горизонтальных террас, какие образует лава на вулканических островах. В таком случае строителю достаточно очень простой инструкции. Но представьте себе, что надо поставить подобный домик на неровном или покатом месте; подумайте, как много дополнительных инструкций должен получить строитель, чтобы суметь в любом случае выполнить свою задачу, несколько меняющуюся от одной площадки к другой. Эта мысленная модель хорошо иллюстрирует, насколько ошибочно дизъюнктивное противопоставление понятий "врожденного" и "выученного" (nature and nurture). Любая способность к обучению основывается на открытых программах, предполагающих не меньше, а больше заложенной в геноме информации, чем так называемые врожденные формы поведения. Тот факт, что это так трудно понять многим в остальном проницательным мыслителям, объясняется, по-видимому, общечеловеческой склонностью мыслить противоположностями.
Адаптивные модификации существуют на всех ступенях органического развития, начиная с самых низших живых существ. Так, например, у многих бактерий, если их выращивают в бедной фосфором среде, разрастаются те химические структуры клетки, которые служат для усвоения этого вещества. Бактериям нужно некоторое время для такого переключения; и если их затем внезапно возвращают в богатую фосфором среду, то они сначала "объедаются" фосфором, пока адаптивная модификация структуры клеток не изменит в обратном направлении их способ питания. Когнитивная функция описанного процесса напоминает здесь функцию регулирующего контура, доставляющего организму информацию о "текущем состоянии рынка".
Описанная выше функция бактерий наводит на мысль о процессе обучения. Но вообще мы называем процессами обучения лишь такие адаптивные модификации, которые относятся к поведению. Получение мгновенной информации, которая не накапливается, т. е. все когнитивные процессы, рассмотренные в предыдущей главе, мы не называем обучением. Отличительным признаком процессов обучения мы считаем то обстоятельство, что некоторое приспособительное изменение происходит в "механике", т. е. в структурах органов чувств и нервной системы,
функцией которых является поведение. Именно в этом изменении структур и состоит приобретение информации, а также ее накопление, поскольку этот процесс более или менее непрерывен.
Адаптирующая модификация, и в особенности адаптирующая модификация поведения, есть когнитивный процесс особого рода. Он превосходит и процесс генома, и все описанные выше процессы приобретения текущей информации в том отношении, что может не только накапливать информацию, как первый из них, но также — как последние — может принимать во внимание и кратковременные изменения окружающей среды. Ни один из рассмотренных выше процессов не способен к тому и другому.
В высшей степени вероятно, что структуры, изменяющиеся при любой адаптивной модификации поведения высших организмов, суть структуры центральной нервной системы. Позже мы еще вернемся к невероятности предположения, будто результаты обучения кодируются в цепных молекулах, наподобие связанной с геномом информации. Чем сложнее живая система, тем менее вероятно, что некоторое случайное изменение ее структуры производит иное действие, чем простая помеха. Но во всем известном нам мире нет более сложной системы, чем устройство высших животных, поведение которых управляется центральной нервной системой. Поэтому весьма удивительно достижение органического становления, выработавшего многообразную адаптивную модифицируемость как раз в этой системе. Эта модифицируемость опирается на удивительные, невероятно сложные структуры, лежащие в основе открытых программ и создающие возможность обучения. Вряд ли в истории человеческого разума было большее заблуждение, чем мнение эмпиристов, будто человек до всякого индивидуального опыта есть чистый лист, "tabula rasa". Впрочем, столь же велико лишь по видимости противоположное, а по существу то же самое заблуждение многих не биологически мыслящих психологов, предполагающих как нечто само собою разумеющееся, что обучение участвует во всех, даже мельчайших элементах поведения животных и человека. Оба заблуждения имеют то вредное следствие, что они заслоняют центральную проблему любого обучения. Проблема эта состоит в следующем: как получается, что обучение улучшает действие поведения, способствующее сохранению вида?
2. СВИДЕТЕЛЬСТВО ЭКСПЕРИМЕНТАЛЬНОЙ ЭМБРИОЛОГИИ
При выполнении открытой программы осуществляется когнитивная, т. е. приспособительная, функция. Внешнее воздействие доставляет информацию, определяющую выбор одной из предусмотренных программой возможностей, и именно той, которая лучше всего подходит к ситуации.
Значительный вклад в понимание этого все еще весьма загадочного процесса внесла механика развития, или экспериментальная эмбриология. Классический пример открытой программы, предлагающей несколько возможностей, доставляет нам эмбриология внешнего клеточного слоя, эктодермы, у эмбрионов позвоночных. В зависимости от места, где
находятся клетки эктодермы в теле зародыша, они могут образовать верхний слой кожи, части глаза или головной мозг со спинным мозгом. Каждая клетка эктодермы содержит информацию, нужную для построения любого из этих органов. Какая из этих программ будет выполнена, зависит от влияний, исходящих от окружения. Если предоставить клетки самим себе, например, в куске, вырезанном из брюшной стороны зародыша лягушки, то эктодерма всегда образует лишь верхний слой кожи. Если же клетка оказывается в близкой окрестности спинной струны, Chorda dorsalis — предшественницы позвоночного столба, — то она образует спинной мозг и головной мозг; там же, где несколько позже к эктодерме приближается выпячивающийся из головного мозга глазной пузырек, она образует точно в надлежащем месте хрусталик. Легко доказать на опыте, что в каждом случае такую специальную форму развития "индуцируют" влияния, исходящие от окрестных образований: если пересадить зародышу лягушки кусочек Chorda dorsalis под кожу живота, то в лежащей над ним эктодерме формируется кусочек нервного
ствола.
Как говорит Шпеман, первый великий исследователь этих процессов, проспект меняя потенция, т. е. первоначально имеющиеся возможности, богаче проспективного значения каждого участка ткани, потому что это последнее в каждом случае зависит от места, где развиваются ее соответствующие куски. Влияния, исходящие от этого места, индуцируют одно из возможных направлений развития, и, когда оно продвинется в достаточной степени, оно окончательно детерминируется, т. е. не может уже быть изменено, так что проспективная потенция соответствующей образующей орган системы далее ограничивается проспектив-ным значением органа.
Все разнообразные виды явлений, происходящих при адаптивных модификациях, по существу, сходны с только что указанными процессами эмбриогенеза. Не очень существенно, исходит ли индуцирующее влияние от окрестности некоторого участка ткани внутри зародыша или от внешнего окружения организма. Система, способная к модификации, всегда содержит генетическую информацию для всех подпрограмм, которые она потенциально способна осуществить. Спинная струна не "говорит", конечно, эктодерме, как она должна сделать спинной и головной мозг, а глазной пузырек не "говорит" ей, как должен выглядеть хрусталик. Поэтому Шпеманово понятие "организатора", с его виталистической окраской, может вызвать некоторое заблуждение. Сейчас мы знаем, что неорганические воздействия также способны "индуцировать", например побудить эктодерму построить тот или иной из упомянутых выше органов. То же относится ко многим адаптивным модификациям, в том числе связанным с поведением. Любое обучение, подобно индукции в механике развития эмбриона, под действием определенных внешних влияний осуществляет ту из разнообразных возможностей открытой программы, которая лучше всего подходит к внешней ситуации. Сами эти внешние воздействия также "предусмотрены", т. е. встроены в программу на основе предшествующих процессов приспособления.
Как жестко и специфично могут быть запрограммированы такие влияния на определенный процесс обучения, видно из опытов X. Гарсиа
и его сотрудников, о которых нам еще придется подробнее говорить впоследствии в другой связи. Здесь достаточно сказать, что крысу нельзя отучить от поедания некоторого вида пищи никакими болезненными наказаниями, за исключением стимулов, вызывающих неприятные ощущения в органах пищеварения.
Итак, обучение в самом широком смысле, определенное как телеономная модификация поведения, по существу, сходно с процессом механики развития, который Шпеман назвал индукцией. (Для читателя, далекого от естествознания, заметим, что "индукцией" называется также метод естественно-научного подхода, который не имеет ничего общего с понятием Шпемана.)
В одном существенном отношении индукция в механике развития отличается от большинства, если не от всех процессов обучения. После того как под действием индукции достигается сужающая детерминация, процесс уже необратим; между тем выученное поведение может быть, как известно, забыто или даже превращено обратной дрессировкой в свою противоположность. В свое время Карл Бюлер серьезно ставил вопрос, не следует ли включить эту обратимость в определение всякого обучения. Примечательно, однако, что есть и такие процессы обучения, которые не обратимы и фиксируются детерминацией в полном шпемановском смысле, раз навсегда. Это, во-первых, процессы так называемого запечатления, необратимо закрепляющие объект определенного инстинктивного поведения, а во-вторых, некоторые процессы, в которых приобретаются интенсивные реакции уклонения, оставляющие, особенно у молодых индивидов, неизгладимый след в виде "психических травм".
Обозначать ли словом "обучение" все телеономные модификации, относящиеся к поведению, — это дело вкуса. Есть авторы, называющие таким образом даже приобретение знаний геномом. И сам я в моей книге "The Innate Bases of Learning"'* соединил под именем обучения все служащие сохранению вида модификации поведения. Поскольку вся психологическая школа бихевиоризма основывает свою работу на гипотезе, что обучение посредством успеха — conditioning by reinforcement — есть единственная форма обучения, более того, единственный заслуживающий изучения процесс во всем поведении животных и человека, мне кажется уместным подчеркнуть своеобразие этого процесса обучения еще и тем, что я рассмотрю его в отдельной главе, а в этой главе описываю лишь простейшие формы индивидуального приобретения знаний.
3. ПРОКЛАДЫВАНИЕ ПУТИ ПОСРЕДСТВОМ УПРАЖНЕНИЯ
Как известно, механизмы автомобиля подвергаются адаптивному изменению с помощью процесса, именуемого "обкаткой". Нечто подобное происходит также, по-видимому, со многими механизмами поведения. Например, М. Уэллс установил, что у только что вылупившихся из яйца каракатиц (Sepia officinalis) реакция поимки добычи уже в первый раз происходит с совершенной координацией, хотя и заметно медленнее,
1 Врожденные основы обучения" (англ.). — Примеч. пер.
чем после многократного повторения. Улучшается также и точность прицела. Э. Гесс наблюдал подобный же эффект упражнения при клевательном движении только что вылупившихся цыплят домашней курицы. Как он показал, попадание в цель не играет никакой роли в улучшении этой формы движения. Гесс надевал цыплятам очки, призматические стекла которых имитировали боковое смещение цели. Цыплята так и не научились корректировать отклонение и все время продолжали клевать в ожидаемом направлении, мимо цели. Но после некоторого упражнения это движение имело гораздо меньший разброс.
4. СЕНСИТИВИЗАЦИЯ
С сенсорной стороны поведения процессам моторного прокладывания путей соответствует так называемая сенситивизация. Так называется вызываемое повторным запуском некоторой реакции снижение пороговых значений запускающих ее ключевых стимулов. Первая реакция вызывает у животного в некотором смысле тревогу; на антропоморфном языке можно было бы сказать, что у него обостряется внимание. Этим сравнением выражается уже и тот факт, что сенситивизация большей частью кратковременное моторного прокладывания путей.
Конечно, состояние тревоги, вызванное сенситивизацией, имеет значение для сохранения вида лишь в тех случаях, когда однократное столкновение с некоторой стимулирующей ситуацией предвещает ее вероятное повторение. Это особенно относится к стимулам, вызывающим бегство. Слегка клюнутый дождевой червь, избежавший гибели благодаря быстрой реакции бегства, правильно поступает, "считаясь" с тем, что опасный дрозд может снова оказаться на его пути. Как показал М. Уэллс, сенситивизация становится особенно важной для сохранения вида, когда объект реакции, враг или добыча, регулярно встречается стаями, как это часто бывает у многих организмов открытого моря. Один из самых впечатляющих примеров сенситивизации поведения поимки добычи — так называемая "feeding frenzy"'* у глубоководных морских рыб, например у акул, макрелей или сельдей. Поймав несколько особей добычи, рыбы кажутся буквально обезумевшими — "frenzy" и означает помешательство — и бессмысленно хватают все вокруг себя, причем пороговые значения ключевых стимулов снижаются настолько, что, например, тунцы хватают толстые крючки без приманки; на этом и основана техника рыболовства, обычная в тропических морях.
У низших животных сенситивизация (Sensitivierung)2* — широко распространенная форма обучения, особенно типичная, согласно М. Уэллсу, для многощетинковых кольчатых червей (Polychaeta). Среди них есть высокоразвитые хищные формы, снабженные эффективными органами чувств.
1 Лихорадка питания" (англ.). — Примеч. пер.
2 Вошедший в употребление термин Sensitierung, обозначающий этот процесс, ошибочен, во всяком случае в немецком языке; в английском же используется слово sensitation.
Как при моторном прокладывании путей, так и при сенситивизации улучшение функции системы достигается самим ее функционированием, что составляет одну из определяющих особенностей обучения. Но в обоих случаях еще отсутствует другой признак, обычно включаемый в определение обучения, — так называемая ассоциация. Этот термин означает образование новой связи между двумя нервными процессами, которые до этого индивидуального процесса обучения функционировали независимо друг от друга. Ассоциация характерна для всех процессов обучения, рассматриваемых в дальнейшем.
5. ПРИВЫКАНИЕ
Стимулирующая ситуация, запускающая при первом столкновении с нею реакцию определенной интенсивности, часто уже во второй раз теряет в некоторой степени свою действенность, а после ряда дальнейших повторений может совсем лишиться запускающей силы. В немецком языке это называется Reizgewohnung (привыканием к стимулу) или Sinnesadaptation (адаптацией к ощущению); как будет видно из дальнейшего, последнее выражение нельзя считать особенно удачным. По-английски это называется "habituation".
В типичном случае исчезновение реакции не зависит от того, следует ли за соответствующим ключевым стимулом дрессирующая, "усиливающая" ситуация стимулирования или нет. Во многом это явление подобно уставанию; может быть, оно и развилось в ходе эволюции из некоторых весьма специфических явлений уставания. Однако его важное значение для сохранения вида состоит как раз в том, что оно предотвращает уставание соответствующей реакции, прежде всего в ее моторном аспекте.
Цель эта достигается тем, что привыкание касается лишь стимулов вполне определенного вида. Пресноводный полип (Hydra) отвечает на целый ряд различных стимулов тем, что стягивает свое тело и щупальца в как можно меньшее пространство. Сотрясение подкладки, прикосновение, небольшое движение окружающей воды, химические или тепловые раздражения — все это производит одно и то же действие. Но если гидра поселяется, как это часто бывает, в медленно текущей воде, где ее тело все время колеблется в разные стороны вследствие завихрений потока, то это стимулирующее действие потока постепенно перестает запускать описанное поведение и полип широко распускает свое тело и щупальца, позволяя им пассивно следовать движению среды. Но при этом — что следует подчеркнуть — пороговое значение всех других стимулов, запускающих стягивание, не меняется. Именно это. без сомнения, произошло бы, если бы стимулы потока не совсем утратили свое действие, а снова и снова вызывали хотя бы очень слабое стягивание гидры. Тогда, конечно, моторная функция реакции уставала бы, а тем самым снижалась бы также способность реагировать на все другие стимулы. Этому и препятствует привыкание к стимулу.
Привыкание можно также назвать десенситивизацией, выработкой нечувствительности. Упомянутое выше выражение "адаптация к ощущению" (Sinnesadaptation), по-английски "sensory adaptation", вводит в за-
блуждение, поскольку его языковая форма вызывает представление, будто имеются в виду процессы, происходящие в органе чувства, подобные адаптации сетчатки нашего глаза к свету и темноте или изменению величины зрачка, служащим для того, чтобы приспособить чувствительность нашего глаза к различным условиям освещения. Конечно, и этот процесс можно называть привыканием; человек, выходящий ночью из ярко освещенной комнаты, может, разумеется, сказать: "Мне надо сначала привыкнуть к темноте". Но в том смысле, как мы понимаем здесь слово "привыкание", оно означает процессы, которые лишь в немногих случаях, как, например, в случае глаза, могут быть сведены к изменениям в самом органе чувства, а происходят в центральной нервной системе. Кроме того, они большею частью долговременнее подлинных "адаптации" органов чувств. Для изучения привыкания к стимулу Маргрет Шлейдт воспользовалась реакцией так называемого клохтания у индюка и доказала, что этот процесс происходит не в органе чувства. Клохтание вызывается разнообразными звуками, и если с помощью генератора звуков производится краткий звук постоянной высоты, повторяющийся через некоторые промежутки времени, то вначале индюк клохчет на каждый из этих стимулов, потом он делает это все реже и наконец совсем перестает. Когда затем производятся звуки другой высоты, то оказывается, что возникшая таким образом десенситивизация относится лишь к очень узкой области высот, примыкающей сверху и снизу к высоте стимулирующего звука. "Кривая адаптации" круто опускается с обеих сторон от вершины, так что пороговые значения высот, чуть дальше отстоящих от исходной, не меняются. Все описанное до сих пор еще укладывается в представление, что адаптация или уставание происходит в самом органе чувства. Но М. Шлейдт показала, что дело обстоит иначе, с помощью опыта, впечатляющего своей простотой: она предложила звук, уже переставший действовать, такой же высоты и длительности, как раньше, но гораздо более тихий. К нашему общему удивлению, этот более тихий звук вновь произвел полное запускающее действие, как если бы был предложен совсем другой звук. Таким образом, специфическая сенситивизация по отношению к данному стимулу, безусловно, произошла не в органе чувства, потому что этот орган в своем адаптированном или уставшем состоянии реагировал бы на тихий звук еще намного слабее, чем на звук прежней силы.
Даже без направленных экспериментов, при наблюдении процессов привыкания в естественных условиях, можно уяснить себе, как сильно исчезновение первоначальной реакции зависит от вполне определенной комбинации внешних стимулов. Уже сама сложность таких условий указывает, что в общем процессе должны участвовать высшие функции Центральной нервной системы. Вот пример: многие из утиных (Anatidae) реагируют на хищников, движущихся по границе их водоема, поведением, которое охотники называют "травлей"*: они преследуют врага, издавая предостерегающие звуки и сколько возможно не упуская его из виду. Эта реакция прежде всего относится к лисе и особенно легко вызывается объектами, покрытыми рыжим мехом, чем весьма коварно пользуются голландские охотники на уток в так называемых загонах*: они привязывают лисью шкуру к спине дрессированной собаки, которая
заманивает уток в длинный, спирально изогнутый канал, так называемую трубку, с ловушкой в конце. Когда мы переселились с нашим богатым поголовьем утиных на озеро Эсс (Ess-See), тогда еще не огороженное от лис, мы опасались, что привычка наших птиц к моим собакам помесям чау и овчарки, с рыжей шерстью и очень похожим на лис может стать для наших птиц опасной. Они подпускали к себе собак столь близко, что подобное поведение по отношению к лисе могло бы оказаться фатальным. Однако это опасение оказалось напрасным, поскольку исчезновение реакции относилось лишь к нашим собственным индивидуальным собакам: даже собака породы чау, принадлежавшая моей знакомой, подверглась "травле" с нисколько не сдерживаемой яростью, и тем более доставалось лисам.
Часто удивляются, насколько малые изменения достаточны, чтобы разрушить привыкание к целой ситуации. Например, достаточно было одной из наших собак появиться на берегу озера, противоположном нашему институту, чтобы вновь разжечь полную реакцию травли у тех же уток и гусей. То же я наблюдал у дроздов шама (Copsychus malabaricus)'*. Пара этих птиц, высиживавших птенцов в моей комнате, изгнала из своего участка птенцов первого выводка, когда следующий выводок приближался к возрасту оперения. Когда я запер в клетку молодого самца, предохранив его таким образом от нападений родителей, прежде всего отца, то взрослые птицы привыкли к присутствию неустранимого сына, так как стимул был некоторым образом скрыт. Они не обращали больше внимания на клетку и ее обитателя. Но когда я неосторожно переставил клетку в другое место комнаты, "адаптация" была совершенно уничтожена и оба родителя так яростно набросились через решетку на молодого самца, что совсем забыли обо всем остальном, и прежде всего о новых птенцах. Поскольку эти птенцы еще не могли самостоятельно есть, они погибли бы с голоду, если бы я не удалил камень преткновения из комнаты.
Явление привыкания задает нам загадки в том отношении, что процесс "адаптации" в ряде случаев кажется явно нецелесообразным. Мы знаем ряд очень специфических форм реагирования, которые, вопреки их очевидному значению для сохранения вида, так скоро десенситивизируются, что, собственно, лишь при первом выполнении проявляют свою полную интенсивность; это показал Роберт Хайнд на реакции предупреждения и бегства, вызываемой у зяблика совой. Даже после нескольких месяцев "отдыха" реакция и отдаленно не достигла той интенсивности, какую она имела в первый раз. Даже сильнейший мыслимый стимул, дрессирующий, т. е. "подкрепляющий", реакцию, а именно преследование живой совой, вырвавшей у зяблика несколько перьев, никоим образом не произвел ожидаемого действия, т. е. не снял притупления этой реакции. Трудно себе представить, чтобы механизм, столь очевидным образом произведенный эволюцией для определенной функции и столь высокодифференцированно выполняющий ее, был создан для того, чтобы развить свою деятельность один раз или самое большее два раза в жизни индивида. Должна быть какая-то ошибка в нашей
1 *Нем. Schamadrosseln. — Примеч. пер
аргументации или в нашей постановке эксперимента. Реакция молодых серых гусей на имитацию родительского предупреждения исчезала в наших опытах так же быстро, как реакция зяблика на сову в опытах Хайнда, и так же мало восстанавливалась. Может быть, мы просто сводим на нет реакцию, запуская ее в человеческом нетерпении слишком быстро и слишком часто; а может быть, мы способствуем ненормально быстрой адаптации тем, что устраиваем в наших "контролируемых" экспериментальных условиях такую однородность всей ситуации, какой никогда не бывает в естественной жизни.
Вольфганг Шлейдт изучил случай, в котором десенситивизация действительно доставляет полезную для приспособления информацию. Как было уже сказано на с. 292, у индюков имеется механизм запуска реакции бегства от хищных птиц, отвечающий очень простой конфигурации стимулов: все, что выделяется черным силуэтом на светлом фоне и движется с определенной угловой скоростью, связанной определенным соотношением с собственной длиной, для дикой индейки является "хищной птицей", например муха, медленно ползущая по белому потолку, точно так же, как пролетающий в небе сарыч*, вертолет или воздушный шар. При попытке сравнить друг с другом разные формы в отношении их действенности, например форму летящего гуся с формой орла, оказалось, что форма, как таковая, вполне безразлична, но привыкание к определенному объекту происходило столь быстро, что в каждом случае действеннее всех оказывался тот, который дольше всего не предъявлялся подопытному животному. На свободе наши дикие индейки проявляли самую сильную "реакцию на хищную птицу", когда показывался дирижабль мюнхенской рекламной фирмы, раз или два в год пролетающий над нашей местностью, гораздо меньшую реакцию — на значительно чаще видимые вертолеты и самую слабую — на сарычей, почти ежедневно круживших над нами. Информация, вызывающая у птицы это быстрое привыкание, в словесном выражении звучала бы таким образом: "Опасайся медленно проходящих по небу предметов, но больше всего тех, которые видишь реже всего ". В естественных условиях Северной Америки это был бы несомненно белоголовый орел (Haliaetus albicilla), единственная хищная птица, которая может угрожать взрослым диким индейкам.
Как уже было сказано, процесс привыкания, или десенситивизации, отличается от ранее рассмотренных простейших процессов модификации поведения — прокладывания путей и сенситивизации — в одном существенном отношении: он сопровождается так называемой ассоциацией, устанавливающей связь врожденных механизмов запуска с весьма сложными функциями распознавания образов, которые будут рассмотрены в одной из следующих глав. Эта связь осуществляет особое, в физиологическом аспекте еще загадочное торможение. В привычной стимулирующей ситуации, которая может представлять невероятно сложную комбинацию отдельных стимулирующих данных, ключевые стимулы, действующие врожденным образом, теряют свое запускающее действие, но сохраняют его во всех других, даже очень мало отличающихся комбинациях с другими стимулами.
6.ПРИУЧЕНИЕ
В обиходной речи мы обозначаем словом "привычка" (Gewohnung) не только процесс, посредством которого мы привыкаем к ранее тягостному стимулу, так что он перестает действовать и более нами не осознается, но и такой процесс, когда определенная стимулирующая ситуация или способ поведения вследствие многократного повторения становятся нам приятными и даже необходимыми. В этом случае, точно так же как и в случае десенситивирующего привыкания, имеется прочная "ассоциация", устанавливающая связь между ключевыми стимулами, действующими на аппарат запуска, и комплексом стимулов окружающей ситуации, повторно сопровождающих такие ключевые стимулы. Эта ассоциация приводит к тому, что реакция, которая первоначально могла быть вызвана простой конфигурацией ключевых стимулов, в дальнейшем нуждается уже для своего запуска во всем комплексе стимулирующих данных, как врожденных, так и "привычных". Таким образом, ассоциация производит здесь действие, прямо противоположное ее действию при десенситивизации, описанному в предыдущем разделе. Там она прекращает действие первоначально действовавших стимулов; между тем в рассматриваемом здесь процессе ключевые стимулы не только остаются действенными в своем взаимодействии с привычной стимулирующей ситуацией, но, более того, их действенность проявляется только в соединении с нею. Значение этого процесса для сохранения вида состоит в значительном усилении избирательности механизма запуска. В отличие от десенситивирующего привыкания, соответствующие примеры можно найти прежде всего у высших животных. Птица, долго содержавшаяся в клетке и годами евшая из одного и того же блюдца, может умереть с голоду, если это блюдце разобьется и ей предложат есть из другой посуды. Патологическим образом приучение проявляется у людей со старческим слабоумием, у которых малейшее изменение обстановки расстраивает осмысленное поведение.
Значение приучения для сохранения вида отчетливее всего видно в онтогенетическом развитии многих животных. Например, только что вылупившийся серый гусь "приветствует", а затем бежит следом за любым предметом, отвечающим на его "свистки покинутости" ритмическими звуками средней высоты и при этом движущимся. Если гусенок проделал это один или несколько раз по отношению к человеку, то в дальнейшем очень трудно побудить его следовать за гусыней или чучелом; а если терпеливо приучить его к этому, он не проявляет уже той интенсивности и верности, какую вызывает у него первый объект. Такая необратимая фиксация побуждения на его объекте, называемая запечатлением, будет рассмотрена дальше, в отдельном разделе. Запечатление в реакции следования гусенка, независимо от того, направлено ли оно на человека или гусыню, вначале относится лишь к виду, а не к индивидуальности запечатленного объекта. Уже способный к бегу и однозначно запечатленный на гусей, маленький гусенок может быть еще без труда перемещен из одной гусиной семьи в другую. Но если он следовал за своими родителями около двух полных дней, то он начинает уверенно
узнавать их индивидуально, и несколько раньше по голосу, чем по чертам лица, — ведь анатиды* примечательным образом узнают друг друга, как и мы, по конфигурации лица. И когда они не видят лица своего собрата по виду, они распознают его еще хуже, чем мы.
Это избирательное привыкание гусенка к индивидуальности своих родителей происходит без участия положительной или отрицательной дрессировки. Случается, что гусенок теряет своих родителей в течение первого часа следования за ними и тогда пытается примкнуть к какой-нибудь другой гусиной паре с выводком, большей частью изгоняющей такого чужака укусами. Но эти неприятные переживания с чужими собратьями по виду не предохраняют его от повторения такой ошибки, а если он снова находит своих родителей, никак не побуждают его крепче их держаться. Напротив, кажется, что даже недолгое следование за чужими гусями стирает образ родителей: как показывают наблюдения, гусенок, однажды потерявший родителей и приставший к чужой паре, склонен снова и снова это повторять. Связанные с этим неприятные переживания, по-видимому, не действуют на его поведение.
Другой пример. Как показал строгими опытами Рене Спите, у человеческого младенца примерно двухмесячного возраста, только что выработавшего моторику улыбки, этот вид приветствия может быть запущен с помощью очень простых макетов. Наряду с конфигурацией двух глаз и переносицы здесь существенно кивающее движение головы, причем оптическое воздействие усиливается отчетливой границей волос. Как добавочный ключевой стимул действует ухмыляющийся рот с высоко оттянутыми вверх уголками. Сначала детский воздушный шар с грубо нарисованными на нем признаками действовал так же, как кивающий воспитатель. Но через несколько недель, в течение которых младенец чаще улыбался подлинным людям, чем макетам, действие простого макета почти внезапно исчезало. Научившись отличать, "как выглядит человек", ребенок боялся теперь разрисованного воздушного шара, которому раньше улыбался, хотя — это следует подчеркнуть — шар не причинил ему никаких неприятных переживаний, так что здесь не могло быть отрицательной дрессировки.
Значительно позже, между шестым и восьмым месяцами жизни, запускающий улыбку механизм еще раз повышает свою избирательность, на этот раз резким скачком. Ребенок начинает, как говорят воспитатели, "дичиться" посторонних, и с этого времени приветствует улыбкой только мать и нескольких других хорошо знакомых людей; по отношению ко всем остальным он заметным образом проявляет поведение бегства или избегания. Вместе с процессом обучения, приводящим к личному узнаванию определенных людей, в ребенке пробуждаются важные процессы образования человеческих связей. Самые ужасные последствия получаются, когда у ребенка отнимают возможность шаг за шагом, как было описано выше, повышать избирательность механизмов запуска своего социального поведения и устанавливать тем самым социальные связи с определенными лицами; между тем это происходит и по сей день в больницах и детских учреждениях, где все время меняется персонал.
Несомненно также, что, когда человеческий младенец "дичится", это происходит вследствие приучения, не связанного с отрицательной дрессировкой, т. е. с неприятными переживаниями от общения с чужими людьми. Напротив, чем меньше чужих видит маленький ребенок, тем сильнее он дичится.
7. РЕАКЦИИ ИЗБЕГАНИЯ, ВЫЗЫВАЕМЫЕ "ТРАВМОЙ"
Я перехожу теперь к процессу обучения, который большинство психологов, изучающих обучение, отождествляют с приобретением подлинного условного рефлекса. Но я полагаю, что здесь идет речь о гораздо более простом явлении, для объяснения которого нет надобности постулировать сложный механизм обратной связи — условные реакции — рассматриваемый в следующей главе.
Ключевой стимул, врожденным образом вызывающий реакцию бегства максимальной интенсивности, часто уже после единственного воздействия неразрывно ассоциируется с сопровождающей его и непосредственно предшествующей ему общей стимулирующей ситуацией. У низших животных этот особый вид ассоциации наблюдается уже на самой низшей ступени. Вероятно, он связан непрерывными переходами с процессами простой сенситивизации. Например, у многих плоских червей световой сигнал, который уже сам по себе, возможно, вызывает незаметную, еще допороговую реакцию бегства, усиливает свое действие при ассоциации с некоторым стимулом, врожденным образом запускающим сильную реакцию бегства; многие американские исследователи поведения называют такое усиление "conditioning"'*. У низших беспозвоночных без централизованной нервной системы приобретение условных реакций — если можно их так назвать — всегда основывается на процессе этого рода. Все их обучение ограничивается этим процессом и видом привыкания, описанным на с. 306 и след. для пресноводных полипов.
У высших животных рассматриваемое здесь приобретение реакций бегства, как и привыкание, ассоциируется с функцией комплексного распознавания образов. Собака, однажды застрявшая во вращающейся двери и испытавшая вследствие этого крайний испуг, с тех пор избегала не только всех вообще вращающихся дверей, но также, очень специальным образом, даже отдаленной окрестности того места, где она пережила травму. Если ей приходилось пробегать по соответствующей улице, то еще до приближения к этому месту она переходила на противоположный тротуар и мчалась мимо него галопом, поджав хвост и опустив уши.
Подобные "психические травмы", как их называют психоаналитики у людей, представляют собой почти необратимые ассоциации между некоторой комплексной стимулирующей ситуацией и реакцией бегства, очень хорошо известные дрессировщикам собак и наездникам: однократное действие стимула на животное может его навсегда "испортить".
1 ""Приведение в требуемое состояние (англ.), термин бихевиористской психологии. — Притч, пер.
8. ЗАПЕЧАТЛЕНИЕ
Необратимая фиксация некоторой реакции на стимулирующей ситуации. встреченной индивидом лишь несколько раз в своей жизни, вызывается также уже упомянутым процессом, который мы назвали запечатлением. С физиологической стороны это явление замечательно тем, что неразрушимая ассоциация формы поведения с ее объектом устанавливается в такое время, когда она еще вовсе не способна проявляться, а в большинстве случаев не может быть обнаружена даже в виде следов. Сенситивный период, в течение которого возможно запечатление, часто располагается в онтогенезе индивида очень рано и в ряде случаев ограничивается немногими часами, но всегда довольно отчетливо определен. Однажды совершившаяся детерминация (с. 303) объекта уже не может быть обращена. Так, например, животные, сексуально запечатленные на другой вид, навсегда и непоправимо "извращены".
Большинство известных процессов запечатления относится к социальным формам поведения. Например, запечатлеваются реакция следования у птенцов выводковых птиц, соперническая борьба у многих птиц, и прежде всего сексуальное поведение. Неправильно говорить, что такая-то птица или такое-то млекопитающее запечатлены, например "запечатлены на человека". То, что таким образом определяется, — это всегда лишь объект некоторой вполне определенной формы поведения. Птица, сексуально фиксированная на чужой вид, может никак не проявлять это в других отношениях, например в отношении сопернической борьбы или иного социального поведения. У серых гусей детские реакции следования и иные социальные формы поведения очень легко запечатлеваются на человека, что весьма полезно для наших исследований; однако сексуальное запечатление при этом не происходит.
Известны также случаи, когда поведение паразитов запечатлевается на вид их хозяина; например, как показал Г. Торп, наездники* откладывают яйца в тот вид гусениц моли, из которого они сами вылупились. Посредством "трансплантации" личинок можно запечатлеть на мучную моль наездников, нормально паразитирующих на восковой моли. У муравьев, как показал Брун, каждый индивид фиксирует свои социальные реакции на том виде муравьев, представители которого оказывали ему помощь, когда он вылупливался из куколки. На этом основывается так называемое рабовладение у многих видов муравьев. Моника Гольцапфель показала, что поведение ловли добычи у сов запечатлевается на определенный вид животных, служащих добычей; более того, если в течение сенситивного периода это запечатление не произошло, то индивид навсегда остается неспособным поймать добычу.
Запечатление связано множеством переходов с другими процессами ассоциативного обучения. Так, например, обучение характерному для вида способу пения у многих певчих птиц, как показал М. Кониси, точно так же связано с некоторым сенситивным периодом и точно так же необратимо, как типичные процессы запечатления. Такие переходы приводили к недоразумениям. Многие авторы, в том числе Р. Хайнд, П. Бейтсон и другие, изучали процессы, вполне отчетливо отличающиеся от типичных процессов запечатления, такие, как процесс присоединения
цыпленка курицы к матери или к некоторому замещающему объекту. Такие явления больше похожи на обычные процессы обучения, чем на типичное запечатление. На основании полученных при этом результатов были подвергнуты сомнению наблюдения Ч. О. Уитмена, О. Гейнрота и мои. Между тем современные результаты К. Иммельмана, М. Шейна, М. Кониси, Ф. Шутца и других полностью подтвердили все установленное прежними авторами уже более двадцати лет назад.
Так же как привыкание и приучение, запечатление "ассоциируется" с комплексными процессами распознавания образов, и так же как в тех двух процессах, при запечатлении выученное "закладывается во врожденный механизм запуска". Тем самым процесс запечатления делает этот механизм более избирательным.
Одна из самых интересных и загадочных функций запечатления состоит в том, что оно осуществляет при восприятии запускающей комбинации стимулов замечательную абстракцию. Сексуальные реакции селезня кряквы, воспитанного в обществе самки пеганки, запечатлеваются не на этот индивид Tadorna tadorna L., а на этот вид. При выборе между многими пеганками этот селезнь почти никогда не выбирает свою "партнершу по запечатлению" — чему препятствуют механизмы, сдерживающие инцест*, — а предпочитает другую представительницу того же вида. Одна галка, воспитанная мною и тем самым "сексуально запечатленная на человека", направила свое токование на маленькую темноволосую девочку. Для меня непостижимо, что побудило птицу считать нас обоих представителями одного вида.
Не решен также вопрос, не играют ли все же некоторую роль в процессе запечатления какие-то вознаграждающие, т. е. дрессирующие, стимулы; иначе говоря, нельзя ли истолковать запечатление как условную реакцию (conditioned response) в смысле И. П. Павлова и американских исследователей психологии обучения. Против этого говорит то уже упомянутое обстоятельство, что запечатленный объект часто оказывается твердо детерминированным к такому моменту времени, когда животное ни разу еще не выполнило, даже в виде слабого намека, относящуюся к этому объекту форму поведения. Например, галка незадолго до вылета из гнезда сексуально запечатлена, хотя можно с уверенностью утверждать, что к этому моменту у нее никогда не было даже намека на сексуальное настроение. Должно пройти еще два года, прежде чем у нее пробудится инстинктивное поведение копуляции, которое, как надо полагать, должно производить важнейшее дрессирующее действие в качестве окончательного акта, удовлетворяющего побуждение. Впрочем, этим не вполне исключается возможное участие других дрессирующих стимулов, еще не распознанных как таковые. Но для такого предположения нет принудительных мотивов, и, по всей вероятности, запечатление есть ассоциативный процесс обучения того же рода, что и оба описанных в предыдущих разделах. Ввиду своей необратимости и своей связи с точно определенными фазами онтогенеза запечатление имеет более отчетливый характер индукции в смысле Шпемана, чем все другие процессы обучения.
9. РЕЗЮМЕ ГЛАВЫ
В предшествующей четвертой главе были рассмотрены физиологические механизмы, принимающие кратковременную информацию и мгновенно оценивающие, но не накапливающие ее. Все они могут функционировать неограниченное число раз, без каких-либо изменений в их механизме от выполнения этой функции. Они составляют основу любого возможного опыта и как раз по этой причине должны быть защищены от любой модификации опытом.
В настоящей пятой главе был рассмотрен принципиально иной процесс, изменяющий в течение индивидуальной жизни самый механизм поведения, и притом таким образом, что улучшается его функция, способствующая сохранению вида.
Улучшение функции некоторой структуры и повышение ее ценности для сохранения вида посредством модификации не более вероятно, чем посредством мутации или рекомбинации наследственных задатков. Но если определенные внешние обстоятельства регулярно вызывают определенные модификации, осуществляющие приспособление именно к этим обстоятельствам, то можно с подавляющей вероятностью предположить, что имеются закрепленные в геноме открытые программы в смысле Эрнста Майра.
Такая генетическая программа содержит несколько отдельных программ для построения некоторого механизма и предполагает поэтому не меньше, а гораздо больше генетической информации, чем единственная замкнутая программа. Но благодаря этому открытая программа способна воспринимать дальнейшую информацию извне, в зависимости от которой определяется, какие из потенциально заложенных в программе возможностей она осуществит. Такое осуществление придает новому приспособлению постоянный характер, так что лежащая в его основе информация накапливается.
Тем самым центральная нервная система повторяет на высшем уровне функцию, уже присущую геному, но отсутствующую у процессов приобретения текущей информации, описанных в четвертой главе.
Любое обучение есть телеономная модификация тех физиологических механизмов, функцией которых является поведение.
Надежную модель открытой программы и адаптивной модификации доставляет нам механика развития, или экспериментальная эмбриология. Какая из "проспективных потенций" некоторой эмбриональной ткани будет осуществлена, зависит от "индуцирующих" влияний окрестности. Все процессы адаптивной модификации, в том числе процессы обучения, по существу, близки к индукции в смысле Шпемана.
Простейшие формы адаптивных модификаций поведения — это прокладывание моторных путей и сенситивизация рецепторных процессов. Последние имеют значение для сохранения вида лишь в тех случаях, когда стимулирующая ситуация с высокой вероятностью появляется в виде серий.
Все дальнейшие процессы модификации, рассмотренные в этой главе, основываются на ассоциации, т. е. на установлении связи между двумя
нервными функциями, ранее не находившимися в причинной зависимости друг от друга. Посредством этого процесса стимулирующая ситуация, часто очень сложная, приобретает влияние на врожденную форму поведения.
В случае привыкания, или десенситивизации, это влияние имеет тормозящий характер: первоначально запускающие ключевые стимулы вследствие ассоциации теряют свою действенность, но сохраняют или соответственно восстанавливают ее даже при малейшем изменении комплексной ситуации. Как знает каждая повариха, "перемена" придает новую действенность притупившимся ключевым стимулам.
При обратном процессе приучения врожденно действующие ключевые стимулы ассоциируются с некоторой комплексной комбинацией стимулов таким образом, что в дальнейшем остаются действенными шшь в их сопровождении. Таким образом врожденный механизм запуска чрезвычайно усиливает свою избирательность.
При интенсивных реакциях бегства запускающие ключевые стимулы часто уже после единственного очень сильного воздействия, именуемого "психической травмой", ассоциируются с сопровождающей комплексной стимулирующей ситуацией, в дальнейшем вызывающей сильное побуждение к бегству. Эта ассоциация часто необратима.
Многие формы поведения, в особенности социального, в ранних сенситивных фазах развития, в которых они еще вовсе не способны функционировать, необратимо фиксируются на некотором объекте. Эти процессы, именуемые запечатлением, своей связью с некоторым сенситивным периодом и своей необратимостью более всех процессов обучения напоминают явления, которые Шпеман назвал индукцией и детерминацией.
В процессах, рассмотренных в четвертом — седьмом разделах этой главы, посредством обучения устанавливается новая связь между независимо функционирующими нервными процессами. Представление об обучении в общем смысле, которого придерживались старые психологи — исследователи поведения, такие, как Вильгельм Вундт и Ч. Л. Халл, весьма точно подходит к описанным процессам. Но если критически сопоставить различные американские теории обучения, как это сделал К. Фоппа в своем прекрасном сжатом обзоре, то снова и снова бросается в глаза, сколько вреда причиняет теоретическим построениям большинства авторов описанная на с. 278—279 тенденция к единообразному объяснению. Снова и снова пытаются справиться со всеми процессами обучения с помощью единственной, всеохватывающей теории. То, что при этом называется "обучением", — это не существующий в действительности промежуточный предмет между процессами, описанными в этой главе, и другими, в основе которых лежит совершенно иная и более сложная организация нервных процессов. Эти процессы составляют предмет следующей главы.
Ваш комментарий о книге
Обратно в раздел психология
|
|


